Концептуализация: как делать и использовать
При погружении в когнитивно-поведенческую психотерапию сразу сталкиваешься с понятием когнитивная концептуализация. Без неё тут никак: собрался на супервизию — неси концептуализацию. Хочешь работать по протоколу — тоже составляй концептуализацию.
В начале практики я думала, что я никогда не пойму, что это такое, как делать концептуализацию, а главное — зачем. Было неясно, как её использовать, и почему я должна тратить время на непонятные упражнения.
TL;DR / Вы прочитаете здесь:
- Когнитивная концептуализация — описание проблемы клиента его словами в схематичном виде
- Концептуализацию можно делать (1) быстро на сессии, (2) на базе когнитивной модели, (3) по общему шаблону по Беку — подробности
- Скачать шаблон концептуализации по Беку для Evernote или Google Doc
- Применение концептуализаций
Концептуализация = описание кейса
Когнитивная концептуализация, простыми словами — диаграмма, которую составляют клиент и терапевт совместно, и которая иллюстрирует психологические проблемы клиента.
Если ещё проще, то концептуализация описывает, ЧТО происходит у клиента, КОГДА, ПОЧЕМУ и ЧТО происходит потом.
Помните, как ослик Иа-Иа? «А всё почему? И по какой причине? И какой из этого следует вывод?»
Важно подчеркнуть 2 момента:
- Клиент и терапевт вместе составляют концептуализацию. Если терапевт самостоятельно составит концептуализацию, теоретически или на основании того, что рассказал клиент, это тоже полезно для понимания, с чем имеешь дело. Но понимание улучшается только для терапевта, на клиента это не распространяется.
- Концептуализация иллюстрирует проблемы клиента — она чётко соотносится с тем, с чем столкнулся клиент. Мы ничего не приписываем и не додумываем. Концептуализация хорошая, если она соответствует опыту клиента, не упускает важные моменты, в ней есть смысл.
Три варианта концептуализаций
Концептуализации можно делать:
- быстрым способом
- на базе когнитивной модели
- по общей схеме, например, по Беку
Быстрый способ — самый простой и понятный. Зарисуйте то, на что жалуется клиент, его собственными словами в виде схемы: что следует за чем и что происходит потом. Обращайте внимание на мысли, эмоции, поведение и последствия. Покажите клиенту и спросите, всё ли так, ничего важное не упущено? Дополните, если надо.
Зарисуйте то, на что жалуется клиент, его собственными словами в виде схемы: что следует за чем и что происходит потом. Обращайте внимание на мысли, эмоции, поведение и последствия. Покажите клиенту и спросите, всё ли так, ничего важное не упущено? Дополните, если надо.
Готово!
Пример быстрой концептуализации
Клиент рассказывает: «Когда я замечаю, что у меня как-то странно бьётся сердце, мне становится страшно. У меня же может быть сердечный приступ! Мне становится ещё страшнее, я пью успокоительные, гуглю симптомы. Через некоторое время проходит, конечно, но…Я хожу по врачам, но никто ничего не может найти, а мне реально плохо. Я стараюсь без лишней необходимости не выходить из дома — вдруг приступ случится, когда я буду на улице. Если так пойдет и дальше, то я буду сидеть дома, не смогу ездить на работу и буду только тратить деньги на врачей.»
Вот быстрая концептуализация словами клиента.
Когнитивная концептуализация — развивающаяся вещь. Со временем станут известны другие факты или обстоятельства. Например, что клиент постоянно сканирует своё тело на предмет, не появились ли симптомы сердечного приступа, или что его дядя по матери умер от сердечного приступа прямо в электричке, а клиенту было тогда всего 6 лет, и это событие произвело на него огромное впечатление. Тогда это можно будет добавить в концептуализацию.
Например, что клиент постоянно сканирует своё тело на предмет, не появились ли симптомы сердечного приступа, или что его дядя по матери умер от сердечного приступа прямо в электричке, а клиенту было тогда всего 6 лет, и это событие произвело на него огромное впечатление. Тогда это можно будет добавить в концептуализацию.
Концептуализировать можно весь кейс в целом, отдельный эпизод или даже отдельную типичную автоматическую мысль. Но это может быть избыточно.
Пример концептуализации на базе когнитивной модели
Мы все знаем, что разработана куча когнитивных моделей, то есть теоретических предположений, как функционирует то или иное расстройство. Например, к той же панике ⏫ когнитивная модель такова:
Держа её в голове, можно точно так же найти ключевые пункты в рассказе клиента и записать их по этой схеме.
Когнитивные модели и порочные круги разработаны для панического расстройства и эпизода паники, для социальной тревоги, для общей тревоги, депрессии, управления гневом, расстройств пищевого поведения и т.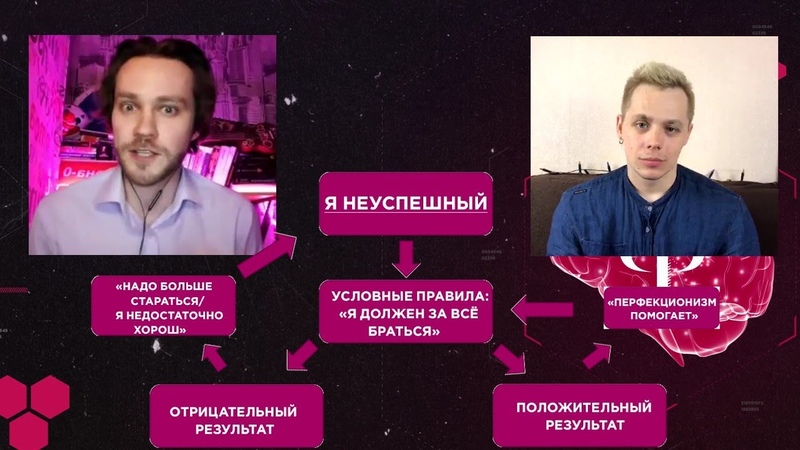 п. Но для неклинических запросов их нет — поэтому используем первый быстрый способ, записывая со слов клиента, что происходит.
п. Но для неклинических запросов их нет — поэтому используем первый быстрый способ, записывая со слов клиента, что происходит.
Пример концептуализации по Беку
И последний вариант когнитивной концептуализации, подойдет под любой случай, но занимает время, да и думать надо. На сессии гораздо лучше использовать быстрый способ.
Значимые события из прошлого или из детства — не значит все события, а только те, которые непосредственно связаны с заявленной проблемой.
Глубинные убеждения — устойчивые представления клиента о себе, например, «Я недостоин любви» или «Я плохой».
Промежуточные убеждения — сопутствующие убеждения и предположения, дополняющие глубинные. Обычно формулируются в связке «если-то» и словами «нужно или должно». Например: «Если тебя кто-то негативно оценивает — это ужасно» или «Если задача слишком трудная, я должен отказаться, и тогда всё будет ок».
Копинг-стратегии — правила поведения, которые помогают клиенту справляться со всей этой историей. Например, «Я перепроверяю всё по несколько раз» или «Я не хожу на вечеринки, где больше 3-х человек народу».
Например, «Я перепроверяю всё по несколько раз» или «Я не хожу на вечеринки, где больше 3-х человек народу».
Дальше расписываем конкретные ситуации, которые могут быть разные.
❗️Для проверки концептуализации взгляните на связанные блоки «Глубинные убеждения» & «Значение автоматической мысли», а также «Копинг-стратегии» & «Поведение» (видите, я их отдельными цветами отметила) — если всё сделано правильно, то между ними есть понятная логическая связь. Если глубинное убеждение никак не соотносится со значением АМ, значит, тут что-то неправильно.
Скачать шаблон когнитивной концептуализации по Беку
Для Google DOC
Для Evernote
Кнопка для донейшена за шаблоны и картинки ⏬
И ещё раз: невозможно с первой встречи с клиентом заполнить концептуализацию на 100%. Она будет уточняться и дополняться, и это правильный путь. Мы возвращаемся и дописываем когнитивную концептуализацию после каждой сессии.
Использование концептуализации
Теперь важный вопрос — и куда её прикладывать, эту концептуализацию?
- Обсуждаем с клиентом.
 Сам факт того, что есть понимание и какая-то схема, уже снимает часть тревог клиента по поводу его проблемы. «О, ну когда вы вот так всё расписали, мне стало уже полегче, и правда, ведь это именно так и работает. А я никогда не смотрел на ситуацию с такой стороны».
Сам факт того, что есть понимание и какая-то схема, уже снимает часть тревог клиента по поводу его проблемы. «О, ну когда вы вот так всё расписали, мне стало уже полегче, и правда, ведь это именно так и работает. А я никогда не смотрел на ситуацию с такой стороны». - Составляем терапевтический план. Под каждый блок в когнитивной концептуализации подбираем соответствующие интервенции.
- Используем на супервизии или интервизии, когда представляем кейс коллегам или супервизору.
- Если кейс оказался трудным, то возвращаемся к концептуализации и смотрим, что мы упустили или в чём ошиблись.
 Сам факт того, что есть понимание и какая-то схема, уже снимает часть тревог клиента по поводу его проблемы. «О, ну когда вы вот так всё расписали, мне стало уже полегче, и правда, ведь это именно так и работает. А я никогда не смотрел на ситуацию с такой стороны».
Сам факт того, что есть понимание и какая-то схема, уже снимает часть тревог клиента по поводу его проблемы. «О, ну когда вы вот так всё расписали, мне стало уже полегче, и правда, ведь это именно так и работает. А я никогда не смотрел на ситуацию с такой стороны».Начинайте с быстрого способа — прямо на сессии записывая словами клиента ЧТО, КОГДА и ПОЧЕМУ происходит, и ЧТО ПОТОМ.
Ну а дальше можно осваивать остальные варианты.
Спасибо, что прочитали. Каждую неделю я отправляю 3 совета, 2 цитаты и 1 вопрос для размышлений на неделе. Подписаны ~ 5000 человек. Присоединяйтесь к нам.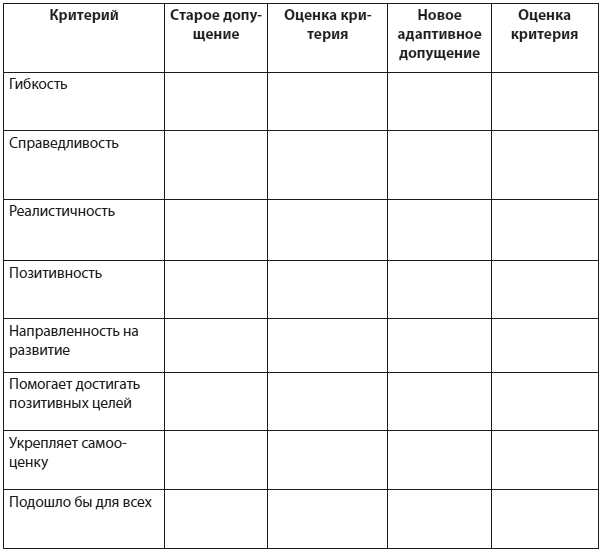
*
ПОДПИСАТЬСЯ
Когнитивная концептуализация и составление плана терапии
Статья подготовлена Васильевой О.Б.
Когнитивная концептуализация
Когнитивная концептуализация обеспечивает терапевту точку отсчета для последующего глубокого понимания клиента. Терапевту следует концептуализировать трудности клиента в когнитивных терминах. Это даст ему возможность определиться в отношении дальнейшего хода терапии, сформулировать цели, успешно работать с автоматическими мыслями, убеждениями или поведением клиента; выбирать подходящие техники и формировать терапевтический альянс. Концептуализация случая необходима для того, чтобы определить наиболее целесообразный и эффективный способ терапии, а также проявить по отношению к клиенту эмпатию – главный компонент, необходимый для установления хороших рабочих отношений.
Главные вопросы, которые должен задать себе терапевт:
«Почему этот человек сидит передо мной?
В чем он уязвим?
Как он компенсировал свои слабости, как справлялся с ними?
Какие жизненные события (травмы, опыт, взаимодействие) оказали на него влияние?
Какие автоматические мысли возникают у него и какими убеждениями они вызваны?»
Другие вопросы, помогающие осуществить концептуализацию, таковы:
Каков диагноз клиента?
Каковы текущие проблемы клиента, как они возникли, за счет чего существуют?
Что привело к развитию расстройства у клиента?
Благодаря каким автоматическим мыслям, образам и поведению существует это расстройство?
Что происходит в жизни клиента в настоящее время и как он это воспринимает?
Каковы типичные ошибки в мышлении этого клиента?
Терапия окажется успешной лишь тогда, когда учтена личная история клиента и система его убеждений, приняты в расчет особенности его восприятия, образ мыслей, эмоций и поведения.
Чтобы проявить по отношению к клиенту эмпатию – не только понять, что он чувствует, но и донести это понимание до клиента, — терапевт должен взглянуть на мир его глазами.
Если относиться к процессу терапии как к путешествию, то концептуализация выполняет функцию дорожной карты. Вначале терапевт и клиент обсуждают цели терапии («пункт назначения»). Туда ведет множество путей и дорог; например, можно двигаться по скоростной трассе или по проселочным ухабистым дорогам. Иногда, столкнувшись с препятствием, приходится менять первоначальный план путешествия и идти в обход. Приобретая опыт в концептуализации, терапевт постепенно наносит на «карту» мелкие детали, делает ее более подробной и точной, чем добивается большей целесообразности и эффективности терапии.
Концептуализация начинается с первой сессии и уточняется (дополняется) на каждой последующей, вплоть до завершения терапии. Основываясь на первоначально предоставленной ему информации, терапевт формулирует гипотезу относительно клиента. Сформулировать гипотезу помогают вопросы:
Сформулировать гипотезу помогают вопросы:
Какие установки, усвоенные в раннем возрасте, пережитый опыт и генетические факторы способствовали возникновению проблемы?
Каковы глубинные убеждения клиента, его промежуточные убеждения и типичные мысли?
За счет каких позитивных и негативных когнитивных, эмоциональных и поведенческих механизмов клиент уравновешивает свои дисфункциональные убеждения?
Как клиент воспринимает (воспринимал) себя, окружающих людей, свой внутренний мир и свое будущее?
Какие стрессовые факторы могли способствовать возникновению его психологических проблем или нарушить его способность разрешить эти проблемы?
На основе новых данных он подтверждает, опровергает или корректирует гипотезу. Концептуализация, таким образом, не является жесткой структурой, она подвижна.
На определенном этапе терапевт представляет свою гипотезу клиенту, и просит оценить ее достоверность. В большинстве случаев, если гипотеза правомерна, клиент подтверждает ее: соглашается, что картина, нарисованная терапевтом, соответствует истинному положению дел.
Концептуализировать проблемы клиента помогает «сведение проблем» или опознание проблем с одинаковыми причинами и их соответствующая группировка. А.Бек приводит в качестве примера женщину с множественными фобиями: лифтов, тоннелей, гор, замкнутых пространств, поездок в открытой машине, полетов на самолете, купания, быстрой ходьбы или бега, сильного ветра и жарких, душных дней. Общее в этих фобиях — страх перед удушьем. Таким образом, в фокусе терапии оказывается следующая мысль: «Я легко могу задохнуться».
Когнитивная концептуализация логически объединяет выявленные у клиента автоматические мысли с более глубокими структурами мышления – убеждениями. В противном случае, если терапевту не удается увидеть более широкую картину, шансы на успех терапии понижаются. Ошибка многих начинающих терапевтов заключается в том, что они часто переключаются с обсуждения одного убеждения на другое, вместо того, чтобы выявить ключевые идеи клиента и последовательно над ними работать. Поэтому, как только терапевт получит данные о типичных автоматических мыслях, эмоциональных реакциях, способах поведения и убеждениях клиента, ему следует начать построение диаграммы когнитивной концептуализации.
Диаграмма когнитивной концептуализации
В диаграмме проблемы клиента представлены в четырех блоках:
1) В первом блоке терапевт приводит три примера ситуаций, вызывающих определенные эмоции или определенное поведение. Здесь описывается сама проблемная ситуация, а также автоматические мысли, значение автоматических мыслей, эмоция и поведение в этой ситуации. Заполняя этот блок, терапевт отвечает на вопросы: Какова проблемная ситуация? О чем клиент думал в проблемной ситуации? Что это значило для него? Какая эмоция сопровождала клиента в этой ситуации? Каковы были действия и поступки клиента?
2) Во втором блоке описываются промежуточные и глубинные убеждения клиента, стоящие за автоматическими мыслями, а также сопутствующие предположения/отношения/правила. Заполняя этот блок, терапевт отвечает на вопросы: Каково ключевые убеждения клиента о себе, других и мире? Каковы предположения, ожидания и правила клиента, т.е. промежуточные убеждения? Как убеждения клиента проявляются в различных жизненных ситуациях и делают его уязвимым, подверженным данному расстройству? Какие промежуточные убеждения помогали клиенту справляться с глубинным убеждением?
3) В третьем блоке терапевт описывает ранний детский опыт, в котором сформировались установки клиента. Заполняя этот блок, терапевт отвечает на вопрос: Как возникло глубинное убеждение и за счет чего оно поддерживалось? Какой жизненный опыт клиента (особенно детский) привел к возникновению и поддержанию данного глубинного убеждения? Каковы в жизни клиента были значимые события, опыт и взаимодействие с окружающими?
Заполняя этот блок, терапевт отвечает на вопрос: Как возникло глубинное убеждение и за счет чего оно поддерживалось? Какой жизненный опыт клиента (особенно детский) привел к возникновению и поддержанию данного глубинного убеждения? Каковы в жизни клиента были значимые события, опыт и взаимодействие с окружающими?
4) В четвертом блоке терапевт описывает компенсаторные стратегии клиента. Обычно главные предположения (убеждения «если…, то…») клиента связывают компенсаторные стратегии клиента с глубинным убеждением. Например: «Если я буду хорошо выглядеть, я буду удерживать мужчин». В этом случае компенсаторная стратегия – забота о своей внешности. Заполняя этот блок, терапевт отвечает на вопрос: Какое поведение клиента помогало ему справиться с глубинным убеждением?
С помощью диаграммы, помимо прочего, можно наглядно увидеть взаимосвязь между глубинными убеждениями, промежуточными убеждениями и текущими автоматическими мыслями клиента. Это своего рода когнитивная карта психопатологии клиента, позволяющая структурировать дальнейшие поступающие от него сведения.
Вначале терапевт заполняет первый блок диаграммы. Терапевт может владеть информацией, позволяющей заполнить только часть диаграммы. В этом случае некоторые поля можно оставить незаполненными или же отметить в них свои предположения, помечая их знаком вопроса, чтобы в дальнейшем проверить их. Получая новые сведения о клиенте, терапевт исправляет или подтверждает свои предположения, постепенно заполняя все поля диаграммы. Стремясь показать клиенту более широкую картину его трудностей, терапевт обсуждает с ним некоторые пункты концептуализации, в некоторых случаях – устно, в других – строит диаграмму вместе с клиентом или же знакомит его с диаграммой, заполненной ранее. Свои собственные предположения терапевт называет гипотезами (или догадками) и просит клиента подтвердить их или опровергнуть.
Диаграмма когнитивной концептуализации должна быть понятна как терапевту, так и клиенту.
Трудности, с которыми клиент сталкивается при выполнении домашних заданий, терапевт использует как возможность улучшить концептуализацию. Он определяет, вызвана ли неудача в выполнении домашнего задания практическими сложностями клиента или его психологической проблемой, которая маскируется под практическую. В этом случае можно предположить перфекционизм (нетерпимость к своим ошибкам), наличие у клиента негативных предсказаний, переоценку усилий, необходимых для выполнения домашних заданий, актуализацию глубинных убеждений при необходимости выполнять домашнее задание.
Он определяет, вызвана ли неудача в выполнении домашнего задания практическими сложностями клиента или его психологической проблемой, которая маскируется под практическую. В этом случае можно предположить перфекционизм (нетерпимость к своим ошибкам), наличие у клиента негативных предсказаний, переоценку усилий, необходимых для выполнения домашних заданий, актуализацию глубинных убеждений при необходимости выполнять домашнее задание.
Составление плана терапии
По ходу терапии психолог постоянно держит в уме вопрос: «Какова проблема клиента и чего я пытаюсь достичь?»
Терапевт стремится добиться ремиссии расстройства и предупредить рецедив, обучить и мотивировать клиента на продолжение самостоятельной работы после завершения терапии.
Терапевт разрабатывает как общий план терапии, так и план каждой сессии.
План терапии основывается на видении клиента терапевтом, на его симптомах и расстройствах, на сформулированных клиентом целях. План терапии помогает составить анализ проблемы клиента, который состоит из описания типичных проблемных ситуаций, дисфункционального поведения, когнитивных искажений и формулировки терапевтических стратегий. План пересматривается и изменяется по мере необходимости согласно индивидуальным потребностям клиента.
План пересматривается и изменяется по мере необходимости согласно индивидуальным потребностям клиента.
Введение в когнитивную терапию Аарона Бека.
Когнитивно-поведенческая терапия. От основ к направлениям
Предисловие. 9
Предисловие научного редактора русского издания. 12
Введение. 15
Глава 1. Введение в когнитивно-поведенческую терапию. 17
Что представляет собой когнитивно-поведенческая терапия?. 18
Какая теория лежит в основе когнитивно-поведенческой терапии?. 19
О чем говорят исследования?. 20
Как развивалась когнитивно-поведенческая терапия Бека?. 22
Каковы основные принципы лечения?. 23
Как проходит терапевтическая сессия?. 29
Как развивается когнитивно-поведенческий терапевт. 30
Как пользоваться этой книгой. 32
Глава 2. Обзор лечения. 36
Выстраивание терапевтических отношений. 36
Планирование лечения и структурирование сессий. 40
Выявление дисфункциональных когниций и реагирование на них.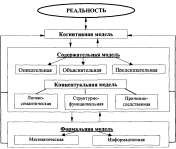 42
42
Акцент на положительном опыте. 46
Содействие когнитивным и поведенческим изменениям
между сессиями (домашние задания). 47
Глава 3. Когнитивная концептуализация. 49
Когнитивная модель. 50
Убеждения. 52
Соотношение поведения и автоматических мыслей. 57
Глава 4. Оценочная сессия. 67
Цели оценочной сессии. 68
Структура оценочной сессии. 69
Как начать оценочную сессию. 69
Фаза оценки. 70
Завершение оценки. 74
Привлечение члена семьи. 74
О ваших впечатлениях. 74
Постановка первоначальных целей лечения и разъяснение
плана лечения. 75
Ожидания от лечения. 77
Разработка исходной когнитивной концептуализации и плана лечения. 79
Глава 5. Структура первой терапевтической сессии. 81
Цели и структура первой терапевтической сессии. 81
Определение повестки дня. 83
Оценка настроения. 84
Оценка изменений. 86
Обсуждение диагноза. 87
Выявление проблем и постановка целей. 91
Знакомство пациента с когнитивной моделью. . . . . . . . . . . . . . . . . . . . . . . . . . .93
. . . . . . . . . . . . . . . . . . . . . . . . . .93
Обсуждение проблемы или поведенческая активация. 97
Подведение итогов сессии и домашнее задание. 97
Обратная связь. 99
Глава 6. Поведенческая активация. 103
Концептуализация бездействия. 103
Концептуализация нехватки ощущения удовлетворения
и удовольствия. 104
Использование графика активности для оценки точности
предсказаний пациентов. 122
Глава 7. Вторая и последующие сессии: структура и формат. 125
Первая часть сессии. 126
Основная часть сессии. 137
Финальное подведение итогов и обратная связь. 144
Третья и последующие сессии. 146
Глава 8. Трудности структурирования терапевтической сессии. 149
Когниции терапевта. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 149
Как перебивать пациента. 150
Социализация пациента. 151
Вовлечение пациента. 151
Укрепление терапевтического союза. 152
Оценка настроения. 153
153
Оценка произошедшего за неделю. 155
Связь между сессиями. 156
Проверка домашнего задания. 159
Обсуждение пунктов повестки дня. 159
Новое домашнее задание. 161
Финальное подведение итогов. 161
Обратная связь. 162
Глава 9. Выявление автоматических мыслей. 163
Характеристики автоматических мыслей. 163
Как объяснить природу автоматических мыслей пациенту. 166
Выявление автоматических мыслей. 168
Обучение пациентов выявлению автоматических мыслей. 181
Глава 10. Выявление эмоций. 184
Отличие автоматических мыслей от эмоций. 185
Трудности в назывании эмоций. 188
Оценка силы эмоции. 190
Использование силы эмоций для планирования терапии. 191
Глава 11. Оценка автоматических мыслей. 193
Выбор ключевых автоматических мыслей. 193
Вопросы, помогающие оценить автоматические мысли. 196
Альтернативные методы оценки мыслей. 204
Когда автоматические мысли оказываются правдой. 209
Обучение пациентов оценке мышления. 211
211
Короткий путь: отказ от вопросов. 212
Глава 12. Реагирование на автоматические мысли. 214
Проверка терапевтических заметок. 214
Оценка и реагирование на новые
автоматические мысли между сессиями. 218
Форма «Проверка автоматических мыслей». 223
Когда заполнение формы не помогает. 223
Другие способы реагирования на автоматические мысли. 225
Глава 13. Выявление и изменение промежуточных убеждений. 226
Когнитивная концептуализация. 226
Модификация убеждений. 241
Глава 14. Выявление и модификация глубинных убеждений. 257
Категоризация глубинных убеждений. 260
Выявление глубинных убеждений. 263
Представление глубинных убеждений пациентам. 264
Информирование пациентов о природе глубинных убеждений
и отслеживание их действия. 265
Создание нового глубинного убеждения. 269
Укрепление нового глубинного убеждения. 270
Модификация негативных глубинных убеждений. 272
Бланк работы с глубинными убеждениями. 272
Глава 15. Дополнительные когнитивные и поведенческие техники. 286
Дополнительные когнитивные и поведенческие техники. 286
Решение проблем и отработка навыков. 286
Принятие решений. 288
Рефокусирование. 291
Оценка настроения и поведения с помощью графика активности. 293
Релаксация и осознанность. 294
Техника последовательного приближения. 295
Экспозиция. 296
Ролевые игры. 297
Техника «пирог». 299
Сравнение с самим собой и списки достижений. 304
Глава 16. Образные представления. 308
Выявление образов. 308
Что рассказывать пациентам об имагинации. 310
Как реагировать на спонтанные образы. 311
Образы как терапевтический инструмент. 320
Глава 17. Домашние задания. 325
Определение домашнего задания. 326
Как повысить вероятность выполнения домашнего задания. 331
Концептуализация трудностей. 340
Проверка домашнего задания. 348
Глава 18. Завершение терапии и профилактика рецидивов. 350
На ранних этапах. 350
В процессе лечения. 351
Перед завершением терапии. 355
Поддерживающие сессии. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 361
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 361
Глава 19. Планирование лечения. 365
Достижение общих терапевтических целей. 365
Планирование лечения между сессиями. 366
Разработка плана лечения. 367
Планирование индивидуальных сессий. 369
Принятие решения об уделении внимания проблеме. 373
Видоизменение стандартного лечения для конкретного расстройства. 379
Глава 20. Проблемы в терапии. 380
Обнаружение проблемы. 380
Концептуализация проблем. 382
Тупиковые ситуации. 391
Решение проблем, возникших на терапии. 392
Глава 21. Развитие когнитивно-поведенческого терапевта. 394
Приложение А. Рабочий бланк терапевтического случая. 398
Приложение Б. Когнитивно-поведенческая терапия: полезные
источники информации. 403
Приложение В. Шкала когнитивной терапии. 405
Список литературы. 412
Общедоступная группа Ассоциация когнитивно-поведенческой психотерапии
КОНЦЕПТУАЛИЗАЦИЯ И РЕСТРУКТУРИЗАЦИЯ ДИСФУНКЦИОНАЛЬНЫХ (ИРРАЦИОНАЛЬНЫХ) УБЕЖДЕНИЙ
Послезавтра, 6 — 7 июня состоится семинар «Концептуализация и реструктуризация дисфункциональных (иррациональных) убеждений» ведущий Д. В. Ковпак.
В. Ковпак.
‼Если вы планируете пройти обучение, то сегодня последний день, когда можно подать регистрацию на участие.
✍🏻Программа семинара:
1. Теоретические предпосылки когнитивной концептуализации.
2. Когнитивно-поведенческие циклы формирования и поддержания расстройств.
3. Классическую модель концептуализации по А. Бек и Дж.Бек.
4 . Современные модификации методов концептуализации.
5. Концептуализацию отдельных видов расстройств:
• Депрессия.
• Тревожные-фобические расстройства (авиафобия, генерализованное тревожное расстройство и др.).
• Паническое расстройство и агорафобия.
📌Развиваемые компетенции:
1. Проводить оценку состояния клиента.
2. Выявлять: автоматические мысли, совпадающие стратегии поведения, промежуточные убеждения, глубинные убеждения.
3. Строить когнитивную концептуализацию.
4. Использовать когнитивную концептуализацию для планирования терапии.
5. Использовать когнитивную концепиуализацию как основу для построения коплайенса.
—————————————————————
📣 Семинар: Концептуализация и реструктуризация дисфункциональных (иррациональных) убеждений.
📅 Дата проведения: 6-7 июня 2020 г.
⏰ Время проведения: с 10:00 до 17:00 (по МСК)
✅ Формат проведения: Online
—————————————————————
👤Ведущий: Ковпак Дмитрий Викторович — кандидат медицинских наук, доцент кафедры психотерапии, клинической психологии и сексологии СЗГМУ им. И. И. Мечникова, член Международного Консультативного Комитета Института Бека (Beck Institute, USA), член правления Исполнительного Совета Международной Ассоциации Когнитивной Психотерапии (IACP), вице-президент Российской Психотерапевтической Ассоциации.
Преподает КПТ с 1998 года, после обучения когнитивно-поведенческой терапии у профессора Г. Кассинова из Института Хофстра, Нью-Йорк, США (H. Kassinove, N.Y., USA). В 2014 году прошел обучение и супервизию у Аллана Мюллера, Д. Судак и других преподавателей Института А. Бека в Пенсильвании, США (Beck Institute for cognitive and behavior therapy, Philadelphia, USA) и многоступенчатый курс обучения РЭПТ в Институте А. Эллиса в Нью-Йорке (А. Ellis Institute, N.Y., USA).
Бека в Пенсильвании, США (Beck Institute for cognitive and behavior therapy, Philadelphia, USA) и многоступенчатый курс обучения РЭПТ в Институте А. Эллиса в Нью-Йорке (А. Ellis Institute, N.Y., USA).
Автор и соавтор более ста научных статей, ряда монографий и научно-популярных изданий. Является председателем Ассоциации Когнитивно-поведенческой психотерапии.
———————————————————-
———————————————————-
𝐏.𝐒: После семинара выдаётся сертификат участника от Ассоциации Когнитивно-Поведенческой Психотерапии.
Репозиторий БГПУ: ИНДИВИДУАЛЬНАЯ КЕЙС-КОНЦЕПТУАЛИЗАЦИЯ В КПТ: ВАРИАНТЫ И ПЕРСПЕКТИВЫ
Пожалуйста, используйте этот идентификатор, чтобы цитировать или ссылаться на этот ресурс:
http://elib.bspu.by/handle/doc/50506
| Название: | ИНДИВИДУАЛЬНАЯ КЕЙС-КОНЦЕПТУАЛИЗАЦИЯ В КПТ: ВАРИАНТЫ И ПЕРСПЕКТИВЫ |
| Другие названия: | INDIVIDUAL CASE CONCEPTUALIZATION IN CBT: OPTIONS AND PROSPECTS |
| Авторы: | Бурдин, М. В. В. |
| Ключевые слова: | издания БГПУ когнитивно-поведенческая терапия номотетическая и идиографическая концептуализация трансдиагностический подход когнитивная модель теории научения теории регуляции эмоций индивидуальная кейс-концептуализация |
| Дата публикации: | 2020 |
| Издатель: | БГПУ |
| Библиографическое описание: | Бурдин, М. В. Индивидуальная кейс-концептуализация в КПТ: варианты и перспективы / М. В. Бурдин // Когнитивно-поведенческий подход в консультировании и психотерапии : сб. ст. II Междунар. науч.-практ. конф., Минск, 5–6 нояб. 2020 г. / Белорус. гос. пед. ун-т ; редкол.: Д. Г. Дьяков [и др.]. – Минск, 2020. – С. 15–21. |
| Краткий осмотр (реферат): | В статье освещаются некоторые проблемы, связанные с преобладанием номотетического типа концептуализации в когнитивно-поведенческой терапии. Представлены существующие методологические способы решения этих проблем, дано описание индивидуальной кейс-концептуализации по J. Persons. Persons. |
| URI (Унифицированный идентификатор ресурса): | http://elib.bspu.by/handle/doc/50506 |
| Располагается в коллекциях: | КОГНИТИВНО-ПОВЕДЕНЧЕСКИЙ ПОДХОД В КОНСУЛЬТИРОВАНИИ И ПСИХОТЕРАПИИ |
Все ресурсы в архиве электронных ресурсов защищены авторским правом, все права сохранены.
✅ Защита кейса на базовом курсе по КПТ – Тома Власова
Защита кейса на базовом курсе по когнитивно-поведенческой психотерапии заключается в следующем:
— проведении, как минимум, 10 консультаций с клиентом, в ходе которых вы записываете либо на видео, либо на аудио ваши сессии;
— создании презентации, где вы описываете все, что было сделано в течение этого времени.
У меня было аж 4 клиента для записи кейса. Изначально я не думала брать так много людей. Конечно, я взяла одного человека, но после 4 консультации стало понятно, что мне этого недостаточно.
Для создания презентации следует составить определенный план, от которого нужно отталкиваться. Обязательно в презентации должны быть:
Обязательно в презентации должны быть:
— концептуализация по Джудит Бек;
— три протокола СМЭР;
— три протокола ЭУМ — это когда мы реструктуризируем наши дисфункциональные эмоции, убеждения во что-то более функциональное и приемлемое;
— план терапии;
— подробный анализ этой работы;
— результаты работы.
Во время написания кейса я столкнулась с такими трудностями:
— при оформлении презентации;
— необходимо было выделить промежуточные убеждения, правила, предположения;
— следовало правильно показать когнитивные искажения.
Моя защита заняла около часа по времени. В моей жизни — это огромный рубеж, потому что тут я уже точно понимаю, что я иду дальше. Я иду на продвинутый курс, который не такой структурированный, как базовый. Теперь я могу сама регулировать процесс обучения и брать те семинары, которые мне самой интересны — это, скорее, клиническая психология и что-то связанное с расстройством личности.
Если у вас есть какие-то вопросы по поводу базового курса, а также, как происходит защита, пишите, пожалуйста, в комментариях. Я с радостью отвечу. #защитакейса #базовыйкурспоКПТ #защитакейсанабазовомкурсепоКПТ
Я с радостью отвечу. #защитакейса #базовыйкурспоКПТ #защитакейсанабазовомкурсепоКПТ
🔸🔸🔸🔸🔸🔸🔸🔸🔸🔸🔸🔸🔸
Консультирую online
Запись в what’s app
+ 7 967 363 83 50
с 10:00 до 20:00
instagram.com/tomavlasova.psy
b17.ru/tamara_vlasova
Пермский центр когнитивной терапии
Клинический психолог, КПТ-терапевт
Диплом с отличием и специализацию «клинический психолог» получила в Пермском государственном национальном исследовательском университете
Прошла долгосрочную программу обучения когнитивно-поведенческой терапии:
• Семинар «Основы когнитивно-поведенческой терапии, КПТ панического расстройства и агорафобии»,16 часов.
• Семинар «Когнитивно-поведенческая терапия депрессии. Концептуализация и работа с убеждениями в КПТ», 16 часов.
• Семинар «Концептуализация в КПТ. КПТ перфекционизма»,16 часов.
• Семинар «КПТ социальной тревожности. Терапевтические отношения в КПТ, позитивная КПТ», 16 часов.
• Семинар «КПТ химических и нехимических зависимостей», 16 часов.
• Семинар «КПТ обсессивно-компульсивного расстройства», 16 часов.
• Семинар «КПТ расстройств пищевого поведения». 16 часов.
• Семинар «КПТ тревоги о здоровье». 16 часов.
• Семинар «КПТ пар. Ошибки и трудности в КПТ». 16 часов.
Прошла программу обучения КПТ детей и подростков, ведущая курса: врач-психотерапевт, к. м. н., международный консультант детского фонда ОСН (ЮНИСЕФ) Марина Веанировна Земляных (Санкт-Петербург):
• «Поведенческая терапия в детской и подростковой практике», 20 часов
• «КПТ при тревожно-фобических расстройствах у детей и подростков», «Использование диалектико-поведенческой терапии в работе с родителями, имеющими детей с эмоциональными нарушениями», 20 часов.
• «КПТ посттравматического стрессового расстройства у детей», «КПТ расстройств пищевого поведения у детей и подростков», 20 часов.
Прошла двухдневный семинар по практике осознанности Mindfulness, ведущая — Онди Уилсон (Великобритания).
Прошла онлайн-курс «CFT и работа со стыдом при тревожных расстройствах, ОКР и дисморфофобии», 12 часов, ведущие — Tara O’Donoghue и Lisa Williams (Великобритания).
Состоит в Ассоциации когнитивно-поведенческой психотерапии (АКПП)
и в Ассоциации когнитивно-бихевиоральных терапевтов (АКБТ).
С июня 2019 года является секретарем Ассоциации когнитивно-бихевиоральных терапевтов (АКБТ), а с декабря 2019 года — руководителем Пермского регионального отделения АКБТ.
Занимается вопросами:
— тревога, панические атаки;
— депрессия;
— неуверенность в себе;
— лень и прокрастинация;
— мотивация к учебе и работе;
— отношения с близкими людьми;
и в особенности:
— страх общения с людьми и публичных выступлений.
функций | Kpt
Цели и спецификации функций
Введение
Config Functions — это клиентские программы, которые упрощают работу на
репозиторий файлов конфигурации Kubernetes.
Примеры использования:
- Проверка конфигурации: например Требовать, чтобы все конфигурации пространства имен
иметь этикеткуМВЗ. - Создание конфигурации: например Предоставить пакет новых услуг
создание пространства именRBAC,
ResourceQuotaи т. Д. - Преобразование конфигурации: например Обновить все
PodSecurityPolicy
конфигурации для улучшения состояния безопасности.

Функции конфигурации
могут выполняться локально или как часть конвейера CI / CD.
В рабочих процессах GitOps функции конфигурации читают и записывают файлы конфигурации из
Репозиторий Git. Изменения в системе, созданные людьми, и изменяющаяся конфигурация
функции проверяются перед передачей на репо. Функции конфигурации могут
запускаться как шаги перед фиксацией или после фиксации для проверки конфигураций перед
они применяются к кластеру.
Почему функции
Мы создаем функции, используя ту же архитектуру, что и остальную часть kpt,
в частности:
- Конфигурация как данные: позволяет нам программно манипулировать
конфигурации с использованием программ без сохранения состояния называются функциями . - Философия Unix: вдохновляет нас на создание каталога полезных,
совместимые функции, которые реализуют
Спецификация функций конфигурации.

Понятия функций
На высоком уровне функцию можно представить так:
-
FUNC: программа, которая выполняет CRUD (создание, чтение, обновление, удаление)
операции на входе.Эта программа может быть упакована как контейнер,
исполняемый файл или скрипт starlark. -
input: Тип списка Kubernetes, содержащий объекты для работы. -
output: Тип списка Kubernetes, содержащий результирующие объекты Kubernetes. -
functionConfig: необязательный объект Kubernetes, используемый для параметризации
поведение функции.
Дополнительные сведения см. В Спецификации функций конфигурации.
Есть две особые функции: функции источника и функции приемника.
Функция источника
Функция источника не требует ввода :
Вместо этого функция обычно производит вывод путем считывания конфигураций
из внешней системы (например, чтение файлов из файловой системы). Найдите примеры в
Найдите примеры в
каталог источников.
Функция раковины
Функция приемника не дает выходного сигнала :
Вместо этого функция обычно записывает конфигурации во внешнюю систему.
(например, запись файлов в файловую систему).Примеры можно найти в каталоге моек.
Трубопровод
Чтобы сделать что-нибудь полезное с функцией, нам нужно составить конвейер
с функцией источника и стока.
Вы также можете использовать оркестратор рабочего процесса на основе контейнера, экспортировав рабочий процесс.
трубопровод. Получите подробные руководства о том, как использовать kpt fn export из
Экспорт руководства по рабочему процессу.
Следующие шаги
Последнее изменение 2 февраля 2021 г .: Добавить действие Github для проверки неработающих ссылок на сайт (# 1393) (8be3f718)
питательных веществ | Бесплатный полнотекстовый | Достижения в средствах, снижающих НАД для лечения рака
6.
 1. Эффективность ингибиторов NAMPT In vivo
1. Эффективность ингибиторов NAMPT In vivo
Кратковременное лечение ингибиторами NAMPT, включая многоцелевые ингибиторы NAMPT или NAMPTi-ADC, продемонстрировало заметную противоопухолевую эффективность на широком спектре моделей опухолей на мышах, либо в виде монотерапии, либо в комбинации. с другими противоопухолевыми препаратами.
Например, FK866 продемонстрировал противоопухолевые, антиангиогенные и антиметастатические эффекты на мышиных моделях рака почки [189]. GNE-617 продемонстрировал сильную противоопухолевую активность на моделях ксенотрансплантатов с дефицитом NAPRT на мышах, включая рак простаты (PC3), фибросаркому (HT1080) и рак поджелудочной железы (MiaPaCa-2), и привел к более чем 98% снижению NAD в опухолях в vivo [190].Аналогичным образом, его структурно родственное производное, GNE-618, продемонстрировало способность подавлять рост опухоли в ксенотрансплантатах NSCLC A549, а также в ксенотрансплантатах рака желудка и саркомы, полученных от пациентов [65,190]. Учитывая, что IDh2-мутантные раковые опухоли полностью зависят от NAMPT для их НАД поставки (поскольку они подавляют экспрессию NAPRT), ингибиторы NAMPT, как было обнаружено, проявляют замечательную противоопухолевую активность против IDh2-мутантных ксенотрансплантатов глиомы и фибросаркомы [53]. В качестве альтернативы, недавнее исследование было направлено на достижение истощения НАД в IDH-мутантных раковых клетках не путем вмешательства в синтез НАД, а путем блокирования регенерации моно-АДФ-рибозных фрагментов из поли (АДФ-рибозы) (PAR) посредством ингибирования поли (АДФ-рибоза) гликогидролаза (PARG), фермент, ответственный за расщепление PAR, с одновременным усилением опосредованного PARP потребления NAD посредством введения агентов, повреждающих ДНК.Как и предполагалось, в модели мыши с мутантной фибросаркомой IDH (HT1080) комбинация темозоломида с нокаутом PARG истощала свободно доступный NAD, предотвращая расщепление PAR, что приводило к гиперпарилированию, секвестрации NAD, метаболическому коллапсу и, действительно, значительному подавлению роста опухоли [191 Несмотря на то, что клеточное старение является противоопухолевым механизмом, так называемый секреторный фенотип, связанный со старением (SASP), предполагает, что стареющие клетки секретируют несколько провоспалительных и проангиогенных факторов и участвуют в создании микроокружения, способствующего развитию опухоли, и способствуют стволу рака.
Учитывая, что IDh2-мутантные раковые опухоли полностью зависят от NAMPT для их НАД поставки (поскольку они подавляют экспрессию NAPRT), ингибиторы NAMPT, как было обнаружено, проявляют замечательную противоопухолевую активность против IDh2-мутантных ксенотрансплантатов глиомы и фибросаркомы [53]. В качестве альтернативы, недавнее исследование было направлено на достижение истощения НАД в IDH-мутантных раковых клетках не путем вмешательства в синтез НАД, а путем блокирования регенерации моно-АДФ-рибозных фрагментов из поли (АДФ-рибозы) (PAR) посредством ингибирования поли (АДФ-рибоза) гликогидролаза (PARG), фермент, ответственный за расщепление PAR, с одновременным усилением опосредованного PARP потребления NAD посредством введения агентов, повреждающих ДНК.Как и предполагалось, в модели мыши с мутантной фибросаркомой IDH (HT1080) комбинация темозоломида с нокаутом PARG истощала свободно доступный NAD, предотвращая расщепление PAR, что приводило к гиперпарилированию, секвестрации NAD, метаболическому коллапсу и, действительно, значительному подавлению роста опухоли [191 Несмотря на то, что клеточное старение является противоопухолевым механизмом, так называемый секреторный фенотип, связанный со старением (SASP), предполагает, что стареющие клетки секретируют несколько провоспалительных и проангиогенных факторов и участвуют в создании микроокружения, способствующего развитию опухоли, и способствуют стволу рака. [192,193].У мышей, несущих ксенотрансплантаты эпителиального рака яичников, добавление FK866 к цисплатину увеличивало выживаемость мышей и задерживало рост обработанных цисплатином опухолей после прекращения лечения [139]. Было показано, что ингибирование NAMPT опосредует это дополнительное противоопухолевое действие за счет i) NAD-опосредованного ингибирования цисплатин-индуцированного SASP при эпителиальном раке яичников и ii) подавления индуцированных платиной раковых стволовых клеток, связанных со старением [139]. Из ингибиторов NAMPT ОТ-82 показал эффективность in vivo против гематологических злокачественных новообразований в зависимости от дозы [109].OT-82 подавлял рост опухоли подкожных ксенотрансплантатов острого миелоидного лейкоза (AML) (MV4-11), эритролейкемии (HEL92.1.7), лимфомы Беркитта (Ramos) и множественной миеломы (RPMI 8226), а также увеличивал выживаемость мышей с системные ксенотрансплантаты ОМЛ (MV4-11), эритролейкоз (HEL92.1.7), ОЛЛ младенческого возраста (MLL-2) и полученные от пациентов ксенотрансплантаты (PDX) ОЛЛ высокого риска [109,154].
[192,193].У мышей, несущих ксенотрансплантаты эпителиального рака яичников, добавление FK866 к цисплатину увеличивало выживаемость мышей и задерживало рост обработанных цисплатином опухолей после прекращения лечения [139]. Было показано, что ингибирование NAMPT опосредует это дополнительное противоопухолевое действие за счет i) NAD-опосредованного ингибирования цисплатин-индуцированного SASP при эпителиальном раке яичников и ii) подавления индуцированных платиной раковых стволовых клеток, связанных со старением [139]. Из ингибиторов NAMPT ОТ-82 показал эффективность in vivo против гематологических злокачественных новообразований в зависимости от дозы [109].OT-82 подавлял рост опухоли подкожных ксенотрансплантатов острого миелоидного лейкоза (AML) (MV4-11), эритролейкемии (HEL92.1.7), лимфомы Беркитта (Ramos) и множественной миеломы (RPMI 8226), а также увеличивал выживаемость мышей с системные ксенотрансплантаты ОМЛ (MV4-11), эритролейкоз (HEL92.1.7), ОЛЛ младенческого возраста (MLL-2) и полученные от пациентов ксенотрансплантаты (PDX) ОЛЛ высокого риска [109,154]. В последней модели было обнаружено, что ОТ-82 задерживает рост лейкемии в 95% (20/21) и вызывает регресс заболевания в 86% (18/21) педиатрических ALL PDX [154].Кроме того, только ОТ-82 продемонстрировал эффективность, сравнимую с химиотерапевтическим режимом индукционного типа, используемым для лечения ОЛЛ у детей, и улучшил эффективность цитарабина и дазатиниба против ОЛЛ у детей у мышей [154]. В соответствии с этими сообщениями, более ранние исследования показали, что FK866 вызывал сильную противоопухолевую активность in vivo в моделях ксенотрансплантата человека ATL, AML, лейкемии Беркитта и лимфомы [173, 174]. Кроме того, в модели ксенотрансплантата лимфомы Беркитта (Ramos) комбинирование FK866 с ритуксимабом привело к увеличению выживаемости мышей и снижению опухолевой нагрузки по сравнению с мышами, получавшими отдельные агенты [148].Кроме того, было показано, что FK866 плюс бортезомиб действует синергетически на моделях ксенотрансплантата множественной миеломы (MM1S) [141]. Также сообщалось, что OT-82 снижает рост опухоли и продлевает выживаемость мышей у мышей с ксенотрансплантатом саркомы Юинга (TC71 и TC32).
В последней модели было обнаружено, что ОТ-82 задерживает рост лейкемии в 95% (20/21) и вызывает регресс заболевания в 86% (18/21) педиатрических ALL PDX [154].Кроме того, только ОТ-82 продемонстрировал эффективность, сравнимую с химиотерапевтическим режимом индукционного типа, используемым для лечения ОЛЛ у детей, и улучшил эффективность цитарабина и дазатиниба против ОЛЛ у детей у мышей [154]. В соответствии с этими сообщениями, более ранние исследования показали, что FK866 вызывал сильную противоопухолевую активность in vivo в моделях ксенотрансплантата человека ATL, AML, лейкемии Беркитта и лимфомы [173, 174]. Кроме того, в модели ксенотрансплантата лимфомы Беркитта (Ramos) комбинирование FK866 с ритуксимабом привело к увеличению выживаемости мышей и снижению опухолевой нагрузки по сравнению с мышами, получавшими отдельные агенты [148].Кроме того, было показано, что FK866 плюс бортезомиб действует синергетически на моделях ксенотрансплантата множественной миеломы (MM1S) [141]. Также сообщалось, что OT-82 снижает рост опухоли и продлевает выживаемость мышей у мышей с ксенотрансплантатом саркомы Юинга (TC71 и TC32). , хотя после прекращения лечения было обнаружено, что опухоли снова вырастают [153]. Примечательно, что комбинация низких доз ОТ-82 и препаратов, усиливающих повреждение ДНК, таких как иринотекан или нирапариб, улучшила противораковую эффективность ОТ-82 в ортотопических ксенотрансплантатах (TC32) и полученных от пациентов ксенотрансплантатах саркомы Юинга [153].Новый ингибитор NAMPT / PAK4, KPT-9274, проявил замечательные противоопухолевые эффекты на широком спектре моделей опухолей у мышей, включая модели почечно-клеточного рака [118], B-клеточного острого лимфобластного лейкоза [194], острого миелоидного лейкоза [169]. , меланома [158], рак толстой кишки [158], тройной отрицательный рак груди [195], протоковая аденокарцинома поджелудочной железы [161], нейроэндокринные опухоли поджелудочной железы [160], рабдомиосаркома [196], саркома Юинга [197], множественная миелома [198] ] и макроглобулинемии Вальденстрема [159].Следует отметить, что противоопухолевая активность KPT-9274 при рабдомиосаркоме, саркоме Юинга, множественной миеломе, раке груди, аденокарциноме протоков поджелудочной железы, карциноме толстой кишки и меланоме в значительной степени определяется ингибированием PAK4 [158,161,195,196,197,198,199]. В целом убедительные результаты доклинических исследований KPT-9274 и OT-82 (суммированные в таблице 3) послужили убедительным обоснованием для оценки этих двух ингибиторов в клинике. Комбинация ингибирования NAMPT с иммунотерапией рака (например.g., антителами против PD1) была выдвинута как многообещающая терапевтическая возможность. Это, в первую очередь, с учетом исследований, показывающих, что когда-то секретируемый во внеклеточную среду, NAMPT способствует поляризации макрофагов в сторону «иммуносупрессивного» фенотипа M2 [200]. У мышей, несущих глиобластому, местная доставка микрочастиц, нагруженных ингибитором NAMPT GMX1778, в сочетании с системной блокадой контрольных точек анти-PD1 приводила к большему привлечению Т-клеток и увеличению выживаемости мышей по сравнению с монотерапией [155].Следует отметить, что одно животное прожило 100 дней и в конце эксперимента было подтверждено отсутствие опухоли. Мыши с опухолями почечной карциномы и меланомы продемонстрировали улучшенный противоопухолевый ответ при одновременном лечении анти-PD1 и KPT-9274 по сравнению с монотерапией анти-PD1 или KPT-9274 [157, 158]. Интересно, что на модели карциномы почек было обнаружено, что монотерапия KPT-9274 значительно снижает уровни NAD + NADH, в то время как комбинированное лечение KPT-9274 и анти-PD1 не привело к общему снижению общих уровней NAD + NADH [157]. .В подтверждение вышеизложенных результатов, повышенная противоопухолевая активность наблюдалась при комбинировании ингибитора NAMPT MV87 и антитела против PD1 по сравнению с отдельными терапиями на модели фибросаркомы [156]. Поразительно, но пополнение NAD посредством добавления экзогенных NMN сенсибилизировало опухоли, устойчивые к терапии анти-PD- (L) 1 (низкий уровень PD-L1), к иммунотерапии на моделях мышей, представляющих устойчивый к иммунотерапии рак легких, рак поджелудочной железы и рак печени [201]. Последний эффект был приписан открытию того факта, что метаболизм NAD управляет индуцированной интерфероном γ (IFNγ) экспрессией PD-L1 в опухолевых клетках через активированную ось NAMPT – TET1 – pSTAT1 – IRF1 – PD-L1, что, в свою очередь, способствует иммунной инвазии опухоли в опухоль. CD8 + T-клеточно-зависимым образом [201].Однако это открытие противоречит другому исследованию, в котором сообщается, что истощение NAD посредством ингибиторов NAMPT усиливает экспрессию PD-L1 в клетках глиобластомы [155]. В целом, эти результаты означают, что необходимо дальнейшее понимание перекрестных помех между NAMPT и PD-L1. Что касается разработки многоцелевых ингибиторов NAMPT, соединения 35 и 7f представляют собой два недавно открытых двойных ингибитора NAMPT / HDAC, которые продемонстрировали превосходное или сопоставимое противоопухолевое действие. эффективность по сравнению с монотерапией FK866 или вориностатом на моделях мышей с ксенотрансплантатом HCT116 [123,124].Совсем недавно Zhang et al. применили методы молекулярной гибридизации с использованием CHS-828 и эрлотиниба в качестве матриц и разработали несколько первоклассных соединений, которые одновременно ингибируют как NAMPT, так и рецептор эпидермального фактора роста (EGFR) [202]. Примечательно, что наиболее активный ингибитор NAMPT / EGFR показал сильное снижение жизнеспособности клеток в нескольких линиях раковых клеток и заметно подавил рост опухоли на мышиной модели рака легкого человека h2975 [202]. Наконец, введение однократной дозы (20 мг / кг) ) двух новых конъюгатов анти-c-KIT антитело-лекарство с новыми ингибиторами NAMPT в качестве полезной нагрузки эффективно блокировали пролиферацию опухолей in vivo в ксенотрансплантатах мышей GIST-T1 стромальной опухоли желудочно-кишечного тракта с положительным c-Kit [117].В соответствии с этими результатами другая группа сообщила о сильной противоопухолевой активности NAMPTi-ADC in vivo в моделях ксенотрансплантатов AML, лимфомы Ходжкина и неходжкинской лимфомы [116].
, хотя после прекращения лечения было обнаружено, что опухоли снова вырастают [153]. Примечательно, что комбинация низких доз ОТ-82 и препаратов, усиливающих повреждение ДНК, таких как иринотекан или нирапариб, улучшила противораковую эффективность ОТ-82 в ортотопических ксенотрансплантатах (TC32) и полученных от пациентов ксенотрансплантатах саркомы Юинга [153].Новый ингибитор NAMPT / PAK4, KPT-9274, проявил замечательные противоопухолевые эффекты на широком спектре моделей опухолей у мышей, включая модели почечно-клеточного рака [118], B-клеточного острого лимфобластного лейкоза [194], острого миелоидного лейкоза [169]. , меланома [158], рак толстой кишки [158], тройной отрицательный рак груди [195], протоковая аденокарцинома поджелудочной железы [161], нейроэндокринные опухоли поджелудочной железы [160], рабдомиосаркома [196], саркома Юинга [197], множественная миелома [198] ] и макроглобулинемии Вальденстрема [159].Следует отметить, что противоопухолевая активность KPT-9274 при рабдомиосаркоме, саркоме Юинга, множественной миеломе, раке груди, аденокарциноме протоков поджелудочной железы, карциноме толстой кишки и меланоме в значительной степени определяется ингибированием PAK4 [158,161,195,196,197,198,199]. В целом убедительные результаты доклинических исследований KPT-9274 и OT-82 (суммированные в таблице 3) послужили убедительным обоснованием для оценки этих двух ингибиторов в клинике. Комбинация ингибирования NAMPT с иммунотерапией рака (например.g., антителами против PD1) была выдвинута как многообещающая терапевтическая возможность. Это, в первую очередь, с учетом исследований, показывающих, что когда-то секретируемый во внеклеточную среду, NAMPT способствует поляризации макрофагов в сторону «иммуносупрессивного» фенотипа M2 [200]. У мышей, несущих глиобластому, местная доставка микрочастиц, нагруженных ингибитором NAMPT GMX1778, в сочетании с системной блокадой контрольных точек анти-PD1 приводила к большему привлечению Т-клеток и увеличению выживаемости мышей по сравнению с монотерапией [155].Следует отметить, что одно животное прожило 100 дней и в конце эксперимента было подтверждено отсутствие опухоли. Мыши с опухолями почечной карциномы и меланомы продемонстрировали улучшенный противоопухолевый ответ при одновременном лечении анти-PD1 и KPT-9274 по сравнению с монотерапией анти-PD1 или KPT-9274 [157, 158]. Интересно, что на модели карциномы почек было обнаружено, что монотерапия KPT-9274 значительно снижает уровни NAD + NADH, в то время как комбинированное лечение KPT-9274 и анти-PD1 не привело к общему снижению общих уровней NAD + NADH [157]. .В подтверждение вышеизложенных результатов, повышенная противоопухолевая активность наблюдалась при комбинировании ингибитора NAMPT MV87 и антитела против PD1 по сравнению с отдельными терапиями на модели фибросаркомы [156]. Поразительно, но пополнение NAD посредством добавления экзогенных NMN сенсибилизировало опухоли, устойчивые к терапии анти-PD- (L) 1 (низкий уровень PD-L1), к иммунотерапии на моделях мышей, представляющих устойчивый к иммунотерапии рак легких, рак поджелудочной железы и рак печени [201]. Последний эффект был приписан открытию того факта, что метаболизм NAD управляет индуцированной интерфероном γ (IFNγ) экспрессией PD-L1 в опухолевых клетках через активированную ось NAMPT – TET1 – pSTAT1 – IRF1 – PD-L1, что, в свою очередь, способствует иммунной инвазии опухоли в опухоль. CD8 + T-клеточно-зависимым образом [201].Однако это открытие противоречит другому исследованию, в котором сообщается, что истощение NAD посредством ингибиторов NAMPT усиливает экспрессию PD-L1 в клетках глиобластомы [155]. В целом, эти результаты означают, что необходимо дальнейшее понимание перекрестных помех между NAMPT и PD-L1. Что касается разработки многоцелевых ингибиторов NAMPT, соединения 35 и 7f представляют собой два недавно открытых двойных ингибитора NAMPT / HDAC, которые продемонстрировали превосходное или сопоставимое противоопухолевое действие. эффективность по сравнению с монотерапией FK866 или вориностатом на моделях мышей с ксенотрансплантатом HCT116 [123,124].Совсем недавно Zhang et al. применили методы молекулярной гибридизации с использованием CHS-828 и эрлотиниба в качестве матриц и разработали несколько первоклассных соединений, которые одновременно ингибируют как NAMPT, так и рецептор эпидермального фактора роста (EGFR) [202]. Примечательно, что наиболее активный ингибитор NAMPT / EGFR показал сильное снижение жизнеспособности клеток в нескольких линиях раковых клеток и заметно подавил рост опухоли на мышиной модели рака легкого человека h2975 [202]. Наконец, введение однократной дозы (20 мг / кг) ) двух новых конъюгатов анти-c-KIT антитело-лекарство с новыми ингибиторами NAMPT в качестве полезной нагрузки эффективно блокировали пролиферацию опухолей in vivo в ксенотрансплантатах мышей GIST-T1 стромальной опухоли желудочно-кишечного тракта с положительным c-Kit [117].В соответствии с этими результатами другая группа сообщила о сильной противоопухолевой активности NAMPTi-ADC in vivo в моделях ксенотрансплантатов AML, лимфомы Ходжкина и неходжкинской лимфомы [116].
6.2. Влияние NA на эффективность ингибиторов NAMPT in vivo
В то время как нормальные ткани экспрессируют NAPRT и могут использовать NA для производства NAD, многие раковые клетки не имеют пути PH для биосинтеза NAD. Следовательно, совместное введение NA с ингибиторами NAMPT было предложено в качестве стратегии для смягчения токсичности ингибиторов NAMPT вне мишени, тем самым расширяя их терапевтический индекс.Согласно этому подходу, экспрессия NAPRT в опухолевых клетках является ключевым фактором, определяющим типы рака, которым он может быть полезен [52]. Действительно, было показано, что добавка NA спасает NAPRT-зависимые линии раковых клеток от NAMPTi-индуцированной цитотоксичности, в то время как она не способна обратить вспять вызванную NAMPTi токсичность в NAPRT-дефицитных клетках [52,53,64,115,118,180,190,203]. с NAPRT-отрицательными опухолями отражена более сложная ситуация. В соответствии с вышеупомянутыми результатами in vitro, ранние исследования показали, что совместное лечение NA устраняет противоопухолевые эффекты FK866, GNE-617 и GMX1777 у мышей, несущих ксенотрансплантаты опухолей, обладающих активностью NAPRT (ML-2 и HCT-116) [190, 203, 204], при этом не влияя на эффективность GMX1777 (24-часовая внутривенная инфузия 150 мг / кг и 650 мг / кг) или LSN3154567 (20 мг / кг; BID) в ксенотрансплантатах раковых клеток с дефицитом NAPRT (HT-1080 и NCI-h2155) [ 115 203].Однако в других экспериментах было обнаружено, что противоопухолевые эффекты in vivo ингибиторов NAMPT, GNE-617 и GNE-618, устраняются добавлением NA даже в ксенотрансплантатах NAPRT-дефицитных клеточных линий и на моделях опухолей, полученных от пациентов [ 190]. Было высказано предположение, что этот спасительный эффект добавленной NA достигается за счет частичного восполнения NAD в опухолях, происходящего независимо от пути PH [190]. В другом исследовании NA обратил антипролиферативный эффект FK866 в NAPRT-дефицитных ксенотрансплантатах A2780 при стандартной дозе 15 мг / кг, а чувствительность к FK866 в присутствии NA была восстановлена при дозе 50 мг / кг, что продемонстрировало даже более значительный противораковый эффект по сравнению со стандартной схемой лечения FK866 (15 мг / кг) [204].Интересно, что в модели множественной миеломы (IM-9) NA аннулировала противоопухолевый эффект GMX1777 (150 мг / кг) при введении сразу после окончания инфузии GMX1777, но когда он вводился через 24 часа после окончания введения GMX1777. инфузии противоопухолевое действие GMX1777 сохранялось [205]. Совсем недавно Шатс и его коллеги сообщили, что зараженные микоплазмами раковые клетки толстой кишки были устойчивы к ингибитору NAMPT STF-118804 в культуре, но также in vivo после ксенотрансплантации голым мышам, и аналогичный эффект наблюдался с E.coli in vitro [25]. Авторы доказали, что раковые клетки защищены от NAMPTi-индуцированного истощения NAD благодаря способности бактерий преобразовывать NAM в NA через свой бактериальный фермент никотинамидазу (PncA) и тем самым поставлять предшественники пути PH [25]. Аналогичным образом, кишечная микробиота, как было продемонстрировано, деамидирует экзогенный NAM в NA и опосредует продукцию NAD в тканях млекопитающих, предполагая, что микробная флора может опосредовать устойчивость опухоли к ингибиторам NAMPT через эндогенное обеспечение NA и участие пути PH [25].Кроме того, было обнаружено, что содержание NA в диете является ключевым фактором, определяющим эффективность ингибитора NAMPT. Например, одно исследование показало, что эффективность ОТ-82 обратно коррелировала с содержанием ниацина (НА) в рационе мышей с лейкемией [109]. Таким образом, сложная системная координация продукции НАД, а также механизмы обмена метаболитов делают так, что эффекты стратегии ингибирования биосинтеза НАД in vivo трудно предвидеть. Кроме того, хотя добавление NA может потенциально улучшить переносимость ингибиторов NAMPT, с этим вмешательством следует проявлять осторожность, поскольку NA может, по крайней мере частично, ослабить их противоопухолевое действие.
6.3. Токсичность ингибиторов NAMPT In vivo
Неблагоприятные эффекты, связанные с ингибиторами NAMPT, были всесторонне исследованы на животных моделях. В остром токсикологическом исследовании лечение мышей высокими дозами FK866 (60 мг / кг 2 раза в день в течение 4 дней) приводило к тромбоцитопении и тяжелой лимфопении, которые купировались совместным лечением NA [204]. В том же исследовании гистологические признаки токсичности, связанной с наркотиками, наблюдались в яичках и селезенке, но не в сетчатке, легких, сердце или головном мозге.Аналогичные гематологические результаты были получены в другом исследовании, которое показало, что крысы, получавшие различные ингибиторы NAMPT, включая FK866 и GM1778, в течение периодов до 15 дней, демонстрировали лимфопению и ретикулоцитопению (снижение количества лимфоцитов и ретикулоцитов), но не тромбоцитопению [206]. -опухолевым мышам инфузия NA снижала смертность, связанную с введением токсичных доз GMX1777 (650 мг / кг и 750 мг / кг) [203, 205]. Точно так же NA защищала мышей B6D2F1 от токсичных доз FK866 [204].Эти результаты подтверждают обоснованность использования NA в качестве антидота для ингибиторов NAMPT и расширения их терапевтического индекса. GNE-617 и GMX1778 были связаны с токсичностью для сетчатки и фатальной сердечной токсичностью в краткосрочных исследованиях безопасности на грызунах, и эти эффекты были воспроизведены в релевантные клеточные системы человека и крысы [97, 207, 208]. Следует отметить, что системное лечение NA не уменьшало токсичность сетчатки, связанную с GNE-617 и GMX1778 у грызунов, и лишь частично защищало их от кардиотоксичности, вызванной NAMPTi (GNE-617) [207,208].Сообщалось также, что ингибиторы NAMPT A-1326133 и A-1293201 вызывают повреждение сетчатки у крыс и собак [209]. С другой стороны, LSN3154567 не вызывал ретинопатию у крыс, получавших 20, 40 и 80 мг / кг этого соединения в течение 4 дней [115]. Тем не менее, токсичность для сетчатки все еще наблюдалась у собак, получавших LSN3154567 (1 и 2,5 мг / кг / день) в течение 4 дней [115]. В отличие от того, что наблюдалось с GMX1778 и GNE-617, совместное введение NA с LSN3154567 (5 мг / кг) действительно защищало собак от ретинопатии [115].В целом, эти результаты предполагают, что повреждение сетчатки, вызванное NAMPTi, и его смягчение с помощью NA могут быть хемотип-специфичными и могут проявлять видовую изменчивость в зависимости от используемой модели на животных. Наконец, недавно сообщенный ингибитор NAMPT ОТ-82 показал благоприятный токсикологический профиль без сердечной, неврологической или ретинальной токсичности у мышей и яванских макак (нечеловеческих приматов), что, по-видимому, позволяет избежать побочных эффектов других ингибиторов NAMPT [109]. Примечательно, что комбинированная терапия низкими дозами ОТ-82 и нирапариба в течение одного месяца привела к нескольким неожиданным смертельным случаям у мышей с саркомой Юинга, что вызвало опасения по поводу потенциальной токсичности, связанной с длительным комбинированным лечением этим соединением [153].Наконец, было показано, что NAMPTi-ADC обладают значительно улучшенным токсикологическим профилем. Например, IgG-8, не связывающийся NAMPTi-ADC, хорошо переносился в дозе 100 мг / кг в исследовании токсичности однократной дозы на крысах и был связан только с легкой острой цитопенией, которая выздоровела через четыре недели после инъекции. без сообщений о тромбоцитопении, токсичности для сетчатки или сердечной недостаточности [116]. Эта доза в 10 раз превышает дозу целевых ADC (αCD19-8 и αCD30-8), при которых обычно проявляется противоопухолевое действие [116].
Использование данных клиентов для повышения эффективности планирования и лечения — Источники для социальных служб
Концептуализация дела: использование данных клиентов для повышения эффективности планирования и лечения
Дениз Р. Вольф, Массачусетс, ATR-BC, ATCS, LPC — 3 CEU
с 9:30 до 12:30
18 марта 2021 г.
Живое интерактивное обучение с использованием платформы Zoom Pro, состоящее из двух частей
Цена на человека $ 89
Описание
Этот трехчасовой интерактивный тренинг познакомит участников со структурированным форматом концептуализации клинического случая.Участники будут использовать этот формат, чтобы уточнить свою гипотезу о потребностях своих клиентов, связанных с уходом и предоставлением услуг, в контексте развития, культурного и межличностного характера. Участники будут работать в небольших группах в комнатах для отдыха, чтобы работать с коллегами над установлением целей совместного лечения с использованием представленного формата концептуализации случая. Будут определены и обсуждены потенциальные индивидуальные, социальные и систематические препятствия на пути к лечению. Участники ознакомятся с этим форматом с целью переноса приобретенных навыков и взглядов на немедленную клиническую реализацию.
Цели обучения:
1. Эффективно разработайте концептуальное представление случая в нескольких форматах для повышения эффективности планирования и лечения
2. Соберите и оцените информацию о факторах развития клиента, семейных, контекстных и защитных факторах
3. Определите факторы развития, ускорения и сохранения, которые способствуют неадаптивное поведение и трудности адаптации
4. Выделите культурные, расовые и гендерные факторы риска и защитные факторы
5.Выявить потенциальные индивидуальные, социальные и системные барьеры, которые часто мешают эффективному лечению
CEU:
Это мероприятие спонсируется Высшей школой социальной работы и социальных исследований Bryn Mawr College (GSSWSR) максимум на 3 кредитных часа. Bryn Mawr College GSSWSR, как школа социальной работы, аккредитованная CSWE, является предварительно утвержденным поставщиком непрерывного образования для социальных работников, профессиональных консультантов и терапевтов по вопросам брака и семьи в Пенсильвании и многих других штатах.
О спикере:
Дениз Р. Вольф, MA, ATR-BC, ATCS, LPC — заместитель директора арт-терапии в Cedar Crest College, а также владелец и практикующий терапевт Mangata Services. Дениз является лицензированным профессиональным консультантом, а также зарегистрированным, сертифицированным советником и сертифицированным супервизором по арт-терапии через Совет по аккредитации арт-терапии. Она получила степень магистра Дрексельского университета. Дениз прошла курс DBT в Вашингтонском университете и смогла учиться непосредственно у основателя DBT Марши Линехан.Более 20 лет Дениз практиковала как терапевт, в основном лечившая подростков с историей сложных межличностных травм. Она работает консультантом во многих местных агентствах и организациях, включая Mastery Charter Schools и Художественный музей Филадельфии, обеспечивая клиническое наблюдение и программы, связанные с оказанием медицинской помощи с учетом травм.
Дениз представляла на городских, региональных, национальных и международных конференциях в области DBT, травматологической помощи, травм и нейробиологии, педагогики и клинического наблюдения.Она опубликовала несколько статей в рецензируемых журналах и написала главы в основополагающие тексты о своей клинической работе с подростками.
Повестка дня
| Время | Содержание | Стиль поставки |
|---|---|---|
| 9:30 — 9:45 | Введение, ориентация на онлайн-семинары. | Ведущий фасилитатора, опытный |
| 9:45 — 10:15 | Концептуализация дела: обоснование и формат | Под руководством фасилитатора |
| 10: 15-10: 45 | Культурное смирение и травма Информированные точки зрения | Ведущий фасилитатора / обсуждение в большой группе |
| 10:45 — 11:15 | Оценка сильных сторон клиентов | Под руководством фасилитатора, экспериментальный |
| 11:15 — 11:30 | ПЕРЕРЫВ | |
| 11:30 — 12 | Работа в малых группах: виньетки концептуализации кейсов | Комнаты отдыха, под руководством участника |
| 12:00 -12: 20 | Отчет и обсуждение в большой группе | Групповое обсуждение, вопросы и ответы |
| 12:20 — 12:30 | Заключение и оценки | Участник под руководством |
Порядок действий : После регистрации вам будет отправлено подтверждение и квитанция об оплате.За несколько дней до тренинга вы получите пакет, содержащий инструкции по входу в сеанс Zoom, раздаточные материалы и требования к вашему сертификату и CEU (если применимо)
Отмена и возврат : Отмена регистрации должна быть произведена по электронной почте ([email protected]) до пяти рабочих дней с даты обучения. Если вы оплатили сбор, вы получите возврат или кредит (на ваш выбор). Если вы не отмените подписку раньше, чем за пять рабочих дней и не заплатите, вы или ваше агентство утратите комиссию.Если вы не отмените обучение в течение пяти рабочих дней с даты обучения и не заплатите, вам или вашему агентству будет выставлен счет на полную оплату. Спасибо
Травматическое повреждение нарушает ядерно-цитоплазматический транспорт и приводит к патологии TDP-43
Существенные изменения:
Как вы увидите из обзоров ниже и из нашего последующего обсуждения, мы были в восторге от этой статьи. Рецензенты задают ряд вопросов, перечисленных ниже, но есть два пункта, по которым мы все согласились, и мы надеемся, что вы сможете решить, возможно, путем включения новых экспериментов или текстовых изменений:
1.В настоящее время вы не демонстрируете напрямую стабильность Nups. Мы предлагаем либо перефразировать текст, либо включить эксперимент с отслеживанием импульсов для изучения стабильности Nups;
Рецензент сделал большое замечание, и мы полностью согласны с тем, что импульсный анализ поможет в уточнении стабильности Nups. Мы подумали о проведении эксперимента по отслеживанию пульса в нашей модели черепно-мозговой травмы (ЧМТ) на мушке. Однако обработка циклогексимидом Drosophila на личиночной или взрослой стадии приводит к летальному исходу (Marco et al., Toxicology Letters, 1982). Более того, посттравматическое повреждение животного становится симптоматическим (Anderson et al., HMG 2018), и мы подозреваем, что лечение их циклогексимидом сделает их фенотипы явно худшими (даже летальными), как сообщалось ранее, что затрудняет выполнение этого анализа in vivo. . Поэтому мы перефразируем наше утверждение о стабильности Nups.
В этом разделе теперь говорится: «Затем мы спросили, изменяет ли травма уровни эндогенного белка нуклеопорина с течением времени. Чтобы решить эту проблему, мы измерили уровни белка Nup214 в мозге травм и контрольных личинок и взрослых особей.Мы провели временной анализ на личиночных (0, 2, 4 и 6 часов) или взрослых (0, 2, 4, 24 и 72 часа) животных после травмы, чтобы оценить уровень белка Nup214 с помощью Вестерн-блоттинга. Интересно, что мы обнаружили, что уровни белка Nup214 остаются повышенными как в мозге личинок, так и в мозге взрослого человека в течение исследуемого периода времени (рис. 1P-S), что позволяет предположить, что травма, вероятно, нарушает уровни Nup214 и, возможно, обмен in vivo ».
2. Ясно обсудите две возможные модели, т.е. агрегирование TDP-43 приводит к изменениям Nup или наоборот; или включить эксперименты, которые проясняют эту взаимосвязь.
Это еще один замечательный момент, сделанный рецензентом 3. В сотрудничестве с доктором Кристофером Доннелли (старший автор) у нас есть рецензируемая рукопись, которая определяет взаимосвязь между агрегацией TDP-43 и секвестрацией Nup62 в двигательных нейронах ИПСК, клетках HEK293T и посмертных тканях пациентов с БАС. Чтобы публично поделиться нашими неопубликованными данными во время проверки, мы загрузили нашу совместную рукопись под названием «Nup62 рекрутируется в патологические конденсаты и способствует нерастворимости TDP-43 в C9orf72 и спорадической БАС / FTLD» на сайте Research Square (Gleixner et al.,). Вот ссылка. https://www.researchsquare.com/article/rs -1 44654 / v1
Вкратце, в нашей совместной рукописи (Gleixner et al., 2021 PREPRINT) мы сообщаем, как TDP43 способствует ядерному истощению и цитоплазматической секвестрации Nup62 в нейронах ИПСК пациентов с БАС / ЛТД. Мы обнаружили, что образование стрессовых гранул, содержащих TDP-43, способствует потере ядер и секвестрации Nup62 in vitro и in vivo. Наши данные свидетельствуют о том, что цитоплазматические взаимодействия Nup62 и TDP-43 являются патологическими по своей природе и приводят к нерастворимым скоплениям как в семейных, так и в спорадических случаях БАС / ЛТД в посмертных тканях мозга (Gleixner et al., Препринт 2021 г.). Что еще более важно, взаимодействие цитоплазматических капель Nup62 через NLS, вероятно, изменяет цитоплазматический фазовый переход TDP-43, приводя к нерастворимым включениям. В дополнение к этому, недавнее исследование показало, что агрегированные и патогенные мутации в TDP-43 способствуют секвестрации и неправильной локализации нуклеопоринов в первичных корковых нейронах мышей, фибробластах человека и индуцированных нейронах, полученных из плюрипотентных стволовых клеток (Chou et al., 2018 Nature Неврология). Важно отметить, что патология ядерных пор наблюдалась в тканях мозга в случаях спорадического БАС и при патогенных мутациях в TDP-43 и C9orf72 (Chou et al., 2018; Кук и др., 2020).
В целом, вышеупомянутые исследования демонстрируют роль TDP-43 в секвестрировании нуклеопоринов при спорадических и семейных формах нейродегенеративных заболеваний человека. Мы включаем подробности об обеих статьях в раздел «Обсуждение» нашей рукописи. В этом разделе обсуждение теперь выглядит следующим образом:
«Недавние данные из нашей лаборатории и лаборатории доктора Доннелли продемонстрировали, что цитоплазматические капли Nup62 проявляют характеристики белков, которые подвергаются фазовому разделению жидкость-жидкость, и вполне вероятно, что это взаимодействие посредством классического NLS способствует вредному фазовому переходу цитоплазматического TDP-43, вызывая он превращается в нерастворимые включения (Gleixner et al., 2021 PREPRINT), обеспечивая механизм того, как Nups могут способствовать патологии TDP-43 при травматическом повреждении. […] Более того, нельзя исключать синергетические эффекты обоих механизмов как причину нейродегенерации при травматическом повреждении ».
Рецензент № 2:
[…] Авторы убедительно демонстрируют, что ЧМТ вызывает морфологические и функциональные изменения в NPC и ядерно-цитоплазматическом транспорте, которые могут быть частично улучшены фармакологическим ингибированием ядерного экспорта, таким образом обеспечивая механистическое понимание потенциального терапевтического пути.При этом есть некоторые аспекты, которые необходимо прояснить, чтобы полностью подтвердить все претензии.
1. На фигуре 1 авторы заключают, что травма увеличивает стабильность Nup214, поскольку наблюдается повышенное накопление Nup214 (посредством WB) со временем посттравматического повреждения. Тем не менее, авторы также сообщают, что уровни мРНК увеличиваются, и предоставленный временной график не позволяет полностью исключить, что повышенные уровни Nup могут происходить преимущественно на уровне транскрипции (а не на уровне белка из-за снижения обмена и / или повышения стабильности, поскольку заявлены авторами).Чтобы обосновать вывод авторов о влиянии повышенной стабильности, следует рассмотреть эксперименты с отслеживанием импульсов и циклогексимидом.
Укрепление таких доказательств стабильности еще более важно, учитывая, что это контрастирует с ранее опубликованными исследованиями, описывающими потерю (или сокращение), а не увеличение компонентов NPC во время нейродегенеративных процессов (Coyne et al., 2020 Neuron; Chou et al. 2017 Nat Neurosc) или старения (D’Angelo et al., 2009 Cell).
См. Наш ответ (1 и 2) на важные комментарии, в которых мы рассмотрели эти проблемы.
2. В своем первоначальном исследовании, описывающем модель TBI Drosophila (Anderson et al. 2018 HMG), авторы предположили, что повторяющаяся травма вызывает неправильную локализацию TDP-43 и образование стрессовых гранул. Здесь авторы предполагают, что патология TDP-43 может быть вызвана TBI-индуцированным дефицитом NPC, как осторожно сказано: «TBI ведет к дефектам NCT, которые потенциально опосредуют патологию TDP-43 при CTE». Учитывая многочисленные доказательства, демонстрирующие, что демиксирование / агрегация TDP-43 индуцирует (in vitro и in vivo) дефицит NPC с возможной секвестрацией некоторых компонентов NPC вместе с агрегатами TDP-43 (Chou et al., 2017 Nat Neuros, Ditsworth et al. 2017 Acta Neuropath, Zhang et al., 2018 Cell, Gasset-Rosa et al. 2019 Neuron), дефицит ядерно-цитоплазматического транспорта, таким образом, может быть следствием цитоплазматической патологии TDP-43, а не причиной. Было бы интересно проверить, спасает ли / предотвращает ли нацеливание на NPC патологию TDP-43, используя siRNA-опосредованное подавление Nups или фармакологическое ингибирование, как это сделали авторы, чтобы показать умеренное улучшение выживаемости или эклозии, соответственно. Предоставление таких доказательств может помочь убедительно определить, вызывает ли дефицит NPC патологию TDP-43 при CTE.
Как было предложено составителем обзора, мы кормили взрослых особей Drosophila KPT-350 (0,05 мМ и 0,5 мМ или ДМСО) в течение 10 дней и обнаружили дозозависимое снижение агрегации Tbph и включили эти новые данные в (дополнительный рисунок 4).
3. Линия 123: было обнаружено, что белки 361 достоверно связаны с ЧМТ. Было ли обнаружено, что они постоянно нарушаются во всех репликах? Следует описать критерии, использованные для отбора этих 361 человека. Это подтвердит их актуальность.
361 ген — это как раз тот, у которых значение p между ЧМТ и не-ЧМТ составляет ≤ 0,05. Когда мы увеличили или уменьшили оба значения> 1,6 раза и p≤0,05, было обнаружено 246 генов, которые были изменены и представляли «протеом TBI». Анализ GO допускал, что все гены изменены в> 1,6 раза и p ≤ 0,1, чтобы получить достаточно генов для проведения анализа GO. Эти гены постоянно наблюдались во всех повторах. Мы включаем эти детали в раздел методологии.
4. Рисунок 2: Сложная модель крысы с черепно-мозговой травмой, где травма, по-видимому, вызывается односторонне и вызывает патологию TDP-43 в ипсилатеральном полушарии, может предоставить уникальную систему для проверки того, распространяются ли агрегаты TDP-43 со временем в нервной системе.Было бы интересно проанализировать контралатеральную сторону и оценить, где патология TDP-43 также в конечном итоге повторяется.
Согласны с рецензентом. Дальнейшие исследования крысиной модели TBI продолжаются, и мы планируем изучить распространение, агрегацию и распространение патологии TDP-43 в различных частях мозга крысы с течением времени. На выполнение и анализ этих анализов на модели грызунов у нас уйдет несколько месяцев. Это выходит за рамки нашей текущей рукописи.
5. На рисунке 3 представлены элегантные доказательства функционального восстановления дефицита NPT с помощью фармакологического ингибирования (с использованием производных KPT-соединения). Улучшилась ли выживаемость или патология TDP-43?
Согласны с рецензентом. Дальнейшие исследования крысиной модели TBI продолжаются, и мы планируем изучить распространение, агрегацию и распространение патологии TDP-43 в различных частях мозга крысы с течением времени. На выполнение и анализ этих анализов на модели грызунов у нас уйдет несколько месяцев.Это выходит за рамки нашей текущей рукописи.
6. Существуют различия между изображениями, показанными на рисунке 2A: не все клетки (окрашенные Dapi) являются Nup-положительными у контрольных мух (в то время как это, по-видимому, имеет место в случае личинок TBI). Что может быть причиной? Представлены ли разные типы клеток в контроле и в ЧМТ?
Все клетки, окрашенные DAPI, являются Nup-положительными на фиг. 2A. Тщательный анализ контрольного изображения на фиг. 2A покажет, что сигналы DAPI окружены окрашиванием Nup, хотя в некоторых клетках сигнал ниже.Это может быть связано с проблемами проникновения антител (техническими). Мы надеемся, что рецензент оценит усилия по получению этих изображений, поскольку трудно получить антитела Nup, которые равномерно окрашивают ткани Drosophila . Более того, даже в пределах одной и той же мозговой ткани Drosophila может быть некоторый уровень вариации в картине окрашивания NPC. Следовательно, мы количественно оцениваем паттерн патологического окрашивания NPC (рис. 2B) в клетках, расположенных конкретно вдоль средней линии VNC, потому что эти клетки показали устойчивое и последовательное окрашивание NPC.
7. Рисунок 2B: Сигнал NPC не согласуется между рисунками 2A и 2B (что выглядит очень пунктирным на последнем), хотя он должен быть похожим: могут ли авторы прояснить это несоответствие?
Мы полагаем, что обозреватель имеет в виду рисунки 2A и 2C. Два изображения отличаются тем, что фигура 2A окрашена только маркером NPC Mab414 (изображение в канале 568), а фигура 2C окрашена как Mab414 (канал 488, зеленый), так и Tbph (канал 568, красный), что, по-видимому, вызывает незначительное изменение сигнала NPC.Мы надеемся, что автор обзора поймет, что, хотя двойное мечение антителами Mab414 и Tbph оказало незначительное влияние на сигнал, аномальная морфология NPC согласована и все еще присутствует на рис. 2B
.
(врезка).
8. Рисунок 6: повышенная интенсивность сигнала ИГХ Nup62, сообщаемая у пациентов с клещевым энцефалитом, не полностью согласуется с данными иммунофлуоресценции. Следовательно, общий фоновый сигнал ИГХ намного выше в тканях пациента по сравнению с показанным единственным контролем.Включение дополнительных средств контроля усилит данные.
Мы включаем дополнительные высококачественные изображения IHC двух элементов управления в нашу исправленную рукопись (см. Дополнительный рисунок 12).
9, Есть проблемы со ссылками. Например: ссылка 45 (строка 121) здесь неверна, это должна быть ссылка 47. Точно так же на странице 134 ссылки, в которых говорится, что «NPC, связанный с нейродегенеративным» цитируются неправильно. Они должны включать Chou et al., 2018; Ditsworth et al. 2017, Zhang et al., Coyne et al., 2020 Neuron, Gasset-Rosa et al., 2019…
Приносим извинения за эту ошибку. Мы тщательно проверили все ссылки и исправили их.
10. Рисунок 2H: основная часть текста относится к стрелкам / наконечникам стрелок, которые должны указывать на аномальное окрашивание RanGAP1, но они отсутствуют на самом рисунке.
Мы добавили стрелки на рисунок. Спасибо, что указали на эту ошибку.
11. Строка 235: цитата неверна. Это должен быть рисунок 3G (а не рисунок 5G).
Мы исправили цитирование рисунка.
Рецензент № 3:
[…] 1. Строка 150 и 414: Здесь авторы заявляют, что они измерили «стабильность белка Nup214». Также в обсуждении они заявляют: «В поддержку наших данных показали, что стабильность и обмен Nup214 нарушаются из-за повторяющихся травм в мозгу мух.грамм. аутофагия). «Однако то, что они делают здесь, — это простой вестерн-блоттинг в разные моменты времени после ЧМТ, и с помощью этой стратегии трудно выявить различия в стабильности белка от синтеза мРНК / белка. Если авторы действительно хотят утверждают, что это изменяет обмен белка, следует выполнить вестерн после остановки трансляции (например, применение циклогексимида), чтобы заявить, что наблюдаемые изменения действительно вызваны изменениями в стабильности белка (не уверен, насколько это возможно в их экспериментальных условиях in vivo. ).В противном случае им нужно провести своего рода эксперимент по маркировке и отслеживанию импульсов. Обратите внимание, что они показывают, что уровень мРНК Nup214 (показан как Nup-214 на Fig1I) действительно увеличивается у взрослых особей и личинок (Fig1I, K). Следовательно, как увеличение транскрипции, так и трансляции или стабильности мРНК могут вносить вклад в увеличение белка Nup214 в дополнение к повышению стабильности белка. Если сложно провести эти эксперименты, чтобы конкретно сказать, что существует дефект в стабильности белка, авторам следует перефразировать свой текст, чтобы сказать, что они наблюдают изменение в «уровнях белка Nup214», и не категорически заявляют, что существует дефект протеостаза Nup214. поскольку у них нет прямых доказательств.
Мы рассмотрели эту проблему более подробно в разделе «Важный комментарий».
(комментарий 1). Мы согласны с рецензентом, и мы переформулируем наше заявление о стабильности белка Nup62 и смягчим его. В частности, мы удалили слово «стабильность» и заменили его на «уровни белка Nup214», как было предложено составителем обзора.
Из-за технологических препятствий мы не можем провести эксперимент с отслеживанием импульсов in vivo. Мы согласны с тем, что уровни мРНК Nup214 также увеличиваются, предполагая, что и TBI влияет на нуклеопорины на уровне транскрипции и трансляции.Мы добавили заявление, в котором рассматриваются обе возможности в обсуждении (см. Ответ на комментарий № 15).
2. Основываясь на модели авторов, TBI вызывает повышенную регуляцию определенных Nups, что, в свою очередь, вызывает агрегацию TDP-43, которая затем вызывает нейродегенерацию. Хотя данные, представленные в этой статье, поддерживают эту модель, также возможно, что TBI сначала вызывает агрегацию TDP-43, которая затем вызывает повышенную регуляцию / агрегацию Nups, что затем приводит к нейродегенерации. Я считаю, что вторая модель здесь не обсуждается и не может быть исключена, поскольку отсутствуют определенные эксперименты, чтобы предпочесть первую модель второй модели.Разница между двумя моделями заключается в том, является ли TDP-43 восходящим или нисходящим потоком от Nup. Например, если сверхэкспрессировать TDP-43 в отсутствие TBI, что происходит с экспрессией и локализацией Nup? Можно ли подавить короткую продолжительность жизни и моторный фенотип, вызванные чрезмерной экспрессией определенного Nup, одновременно подавляя TDP-43? Возможно, обе модели 1 и 2 верны, и в этом случае это может означать, что TDP-43 и Nups могут регулировать друг друга, вызывая порочный круг в патогенезе ЧМТ.
Рецензент высказал обоснованное мнение. Мы остаемся непредвзятыми и открытыми в отношении обеих моделей.
Ранее сообщалось, что сверхэкспрессия самого C-концевого фрагмента TDP-43 (TDP43-CTF) в первичных корковых нейронах нарушает локализацию и распределение Nup98. Это также очевидно из окрашивания mAb414, показывающего, что сверхэкспрессии TDP43-CTF достаточно, чтобы вызвать патологию FG Nup (Chou et al., 2018). Эти находки предполагают каскад событий, вызванных дефектами ядерно-цитоплазматических транспортных процессов.Мы планируем продолжить эксперименты, которые автор обзора предложил, чтобы напрямую оценить, какие механизмы задействованы и какая модель более актуальна для нейродегенерации, опосредованной TBI. Мы включаем утверждения об обеих моделях в раздел «Обсуждение» (см. Ответ на важный комментарий №2).
https://doi.org/10.7554/eLife.67587.sa2
Количественная оценка силы перекрестных помех от кворума внутри микробных сообществ
Abstract
В многовидовых микробных сообществах обмен сигналами, такими как лактоны ацил-гомосерина (AHL), позволяет общаться внутри и между видами грамотрицательных бактерий.Этот процесс, широко известный как определение кворума, помогает в регулировании генов, имеющих решающее значение для выживания видов в гетерогенных популяциях микробов. Хотя обмен сигналами широко изучался в хорошо перемешанных средах, о последствиях перекрестных помех в пространственно распределенных смесях видов известно меньше. Здесь динамика передачи сигналов была измерена в пространственно распределенной системе, содержащей несколько штаммов, использующих гомологичные системы передачи сигналов. Перекрестные помехи между штаммами, содержащими цепи AHL-рецепторов lux , las и rhl , были определены количественно.В распределенной популяции микробов влияние состава сообщества на пространственно-временную динамику было охарактеризовано и сравнено с результатами моделирования с использованием модифицированной модели реакции-диффузии. После введения одного члена для учета перекрестных помех между каждой парой сигналов модель смогла воспроизвести паттерны активации, наблюдаемые в экспериментах. Мы количественно оценили устойчивость распространения сигнала в присутствии взаимодействующих сигналов, обнаружив, что динамика передачи сигналов в значительной степени устойчива к помехам.Способность нескольких диких изолятов участвовать в AHL-опосредованной передаче сигналов была исследована, выявив различные сигнатуры перекрестных помех для каждого вида. Наши результаты представляют собой способ охарактеризовать перекрестные помехи между видами и предсказать динамику передачи сигналов на системном уровне в многовидовых сообществах.
Информация об авторе
В природе бактерии обычно встречаются в пространственно неоднородных смесях. В этих средах несколько видов общаются с помощью химических сигналов, и перекрестные помехи часто регулируют деятельность микробных популяций, включая взаимодействие с системой хозяина, формирование биопленок и биолюминесценцию.Понимание таких бактериальных взаимодействий важно для контроля и предотвращения такого поведения на популяционном уровне, регулируемого обменом сигналами. Кроме того, количественная оценка бактериальных перекрестных помех поможет повысить надежность синтетических сотовых сетей, использующих обмен сигналами.
Хотя сотовая сигнализация понимается в хорошо смешанных системах с одним сигналом, нам не хватает детального понимания сигнализации в пространственно распределенных сотовых сетях или сетях с несколькими сигналами. Мы создали экспериментальную систему для наблюдения и количественной оценки микробного взаимодействия между тремя бактериальными языками.Была реализована математическая модель для прогнозирования последствий обмена множественными сигналами в сотовых сетях, и было обнаружено хорошее согласие между экспериментальными результатами и теоретическими предсказаниями. В математической модели одного параметра было достаточно для учета перекрестных помех между видами бактерий. Эти экспериментальные и теоретические инструменты позволяют нам лучше понять и предсказать, как передача сигналов влияет на поведение как естественных, так и синтетических микробных сообществ.
Образец цитирования: Silva KPT, Chellamuthu P, Boedicker JQ (2017) Количественная оценка силы перекрестных помех, обнаруживаемых кворумом в микробных сообществах. PLoS Comput Biol 13 (10):
e1005809.
https://doi.org/10.1371/journal.pcbi.1005809
Редактор: Олег А. Игошин, Университет Райса, США
Поступила: 21 апреля 2017 г .; Принят к печати: 5 октября 2017 г .; Опубликован: 19 октября 2017 г.
Авторские права: © 2017 Silva et al.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Все соответствующие данные находятся в документе и его файлах с вспомогательной информацией.
Финансирование: Эта работа была поддержана премией Управления военно-морских исследований номер N00014-15-1-2573 (https://www.onr.navy.mil/) и премией молодых факультетов Агентства перспективных исследовательских проектов Министерства обороны США D16AP00121 ( https: // www.darpa.mil/). Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Микробы общаются друг с другом, чтобы координировать поведение и экспрессию генов посредством процесса, известного как определение кворума. Некоторые грамотрицательные бактерии используют ацил-гомосериновые лактоны (AHL) в качестве сигнала для коммуникации [1–6].Эти сигнальные системы обычно состоят из синтазы, такой как luxI , которая продуцирует вариант AHL, и рецептора, такого как luxR , который связывается с AHL. Рецептор вызывает глобальные изменения экспрессии генов в ответ на высокие концентрации AHL. Было охарактеризовано более 150 систем восприятия кворума [7,8], причем большинство видов содержат один или несколько сигнальных путей. Каждая система обычно производит одну доминирующую версию AHL [7,8], и на сегодняшний день идентифицировано 56 различных AHL [7].Варианты AHL включают изменения длины углеродной цепи, идущей от лактонового кольца, и химические модификации этой углеродной цепи, такие как добавление карбонильных групп [2,7]. Вариация химической структуры AHL влияет как на сродство к рецептору, так и на регуляторный ответ [6–8].
Сообщалось о нескольких примерах перекрестных помех между сигнальными микробами, при которых сигнал, производимый одним видом, связывается с рецептором другого вида [1,6,9–14].Например, Chromobacterium violaceum , патогенная грамотрицательная бактерия, которая продуцирует более короткие цепи AHL, активирует экспрессию генов в Vibrio harveyi , грамотрицательных морских бактериях, которые продуцируют более длинные цепи AHL [9]. Каждая такая пара AHL и рецептора в различной степени ингибирует или способствует активации экспрессии гена. Многовидовые сообщества коллективно производят сложные смеси сигналов, и на активацию экспрессии генов в сообществе влияют перекрестные помехи между различными вариантами AHL.Количественная оценка AHL-опосредованных перекрестных помех поможет нам построить прогнозное понимание динамики передачи сигналов в гетерогенных микробных популяциях, потенциально позволяя нам контролировать активацию экспрессии генов в естественных и синтетических микробных сообществах [1,2,5,9,15-19] .
Здесь мы разрабатываем и внедряем анализ передачи сигналов с использованием штаммов отправителя, получателя и интерактора для измерения динамики передачи сигналов в популяциях, содержащих несколько AHL, см. Рис. 1A. Этот анализ перекрестных помех оценивает надежность обмена сигналами в смешанных микробных сообществах.Здесь мы используем штамм-отправитель, продуцирующий AHL 3-оксо-C6 HSL, и множественные штаммы взаимодействия, продуцирующие сигналы, включая 3-оксо-C12 HSL, производимый синтазой LasI и C4-HSL, продуцируемый синтазой RhlI. Наша работа расширяет объем предыдущих исследований, которые были сосредоточены на обмене сигналами в хорошо перемешанных средах, где диффузия AHL играет минимальную роль [12,17,18,20–23]. Первоначальная работа выполнена Кантоном и др. [20] количественно исследовали способность нескольких AHL связываться с одним рецептором в планшет-ридере. Wu et al.[23] использовали микрофлюидный подход и метод проточной цитометрии для измерения взаимодействий между контурами люкс и las . Другие исследования, такие как McClean et al. [11] наблюдали ответ сигнальной системы в Chromoacterium violaceum на очищенные варианты AHL. Хотя диффузия играет здесь роль, не было проведено значительной количественной работы, чтобы уловить пространственные эффекты. В другом исследовании Dilanji et al. [24] изучили влияние диффузного фронта волны АГЛ, создаваемого экзогенно добавленным химическим веществом на пластинах с агаром.В качестве альтернативы внешнему добавлению AHL мы добавили отправителя E . Колония coli к середине планшета, способная синтезировать молекулы AHL, как показано на рис. 1B и аналогично предыдущим экспериментам [25]. Наш анализ включает в себя штамм интерактора для определения устойчивости передачи сигналов AHL к перекрестным помехам от нескольких сигналов AHL, включая сигналы, продуцируемые изолятами диких бактерий.
Рис. 1. Экспериментальный анализ для количественной оценки перекрестных помех между сигналами восприятия кворума бактерий.
A. Анализ перекрестных помех измеряет последствия внесения переменного количества штамма взаимодействия в сеть контроля кворума, содержащую штаммы отправителя и получателя. B. Штамм-отправитель помещается в середину агаровой подушки, окруженный однородной смесью штаммов-получателей и взаимодействующих штаммов. C. Когда сигнал от колонии-отправителя распространяется наружу, штамм-получатель вырабатывает красный флуоресцентный белок в ответ на пороговый уровень сигнала. D. Сравнивается пространственная динамика экспрессии генов в приемниках при наличии и отсутствии штамма интерактора. Гипотетические кривые показывают, как штамм интерактора сдвигает кривую активации, либо способствуя активации (возбуждающие перекрестные помехи), либо подавляя активацию (ингибирующие перекрестные помехи).
https://doi.org/10.1371/journal.pcbi.1005809.g001
Сообщалось об обмене несколькими AHL [14,26], но эти исследования минимизировали перекрестные помехи за счет использования AHL, которые будут слабо взаимодействовать друг с другом.Следовательно, перекрестные помехи AHL в естественных условиях, когда множественные сигналы обмениваются в пространственно структурированных сообществах, не совсем понятны. Наше исследование экспериментально измерило влияние напряжения соседнего интерактора на временную шкалу передачи сигналов между деформациями отправителя и получателя. Штамм интерактора производит неродственный сигнал AHL, который влияет на способность штамма-получателя реагировать на родственный сигнал AHL, излучаемый штаммом-отправителем. Последствия таких перекрестных помех были изучены на масштабах, намного превышающих размеры отдельных ячеек.Была получена математическая модель и проведено сравнение с экспериментальными результатами для прогнозирования сложных взаимодействий с рецептором AHL, происходящих в популяциях микробов в природе.
Результаты
Обнаружение перекрестных помех при определении кворума с использованием установки для анализа на планшете
Наша экспериментальная установка была основана на анализе планшета типа отправитель-приемник LuxI / LuxR [14,20,24,25,27,28] с добавлением взаимодействующего штамма. В этой установке штамм-отправитель продуцирует AHL 3-oxo-C6 HSL [21], который затем связывается с рецепторным белком LuxR и активирует экспрессию генов в отправителях, когда присутствует достаточное количество AHL.Экспрессия активированного гена в отправителях приводит к увеличению продукции GFP, см. Рис. 1A. Ресиверы обладают способностью продуцировать белок рецептора LuxR, и в присутствии достаточного количества AHL они активируют экспрессию гена RFP. Интераторы конститутивно продуцируют неконфликтный AHL, соответствующий генам синтазы lasI или rhlI . Плазмиды, кодирующие эти конструкции, показаны на рисунке A в файле S1. Совместное культивирование получателя и отправителя привело к трехкратному увеличению флуоресценции, а введение интеракторов привело к эквивалентному или уменьшенному уровню флуоресценции, см. Рисунок B в файле S1.
Клетки-приемники, содержащие регулируемый LuxR флуоресцентный репортер, распределяли на агаровой подушке LB диаметром 20 мм, содержащей колонию штамма-отправителя в середине планшета, см. Фиг. 1В. Отправитель содержит плазмиду, экспрессирующую цепь LuxRI, см. Рисунок A в файле S1. Время активации экспрессии гена в приемниках измеряли в зависимости от расстояния от колонии-отправителя. Добавляя взаимодействующий штамм, производящий дополнительный AHL, в лужайку ячеек-получателей, сдвиг в профиле активации количественно определяет перекрестные помехи между штаммом-взаимодействующим и системой отправитель / получатель.
Как показано на рис. 1C, в ячейках приемника для анализа планшета, смежных с отправителями, экспрессия RFP составляет около 5,25 часа, а клетки-приемники, расположенные на больших расстояниях, активировали RFP через 6–8 часов. Для количественной оценки распространения активации внутри планшета использовались флуоресцентные изображения RFP для расчета времени, необходимого для активации экспрессии RFP в приемниках на нескольких расстояниях от колонии-отправителя. Активация RFP определяется как когда по крайней мере 10% пикселей, принадлежащих ячейкам, имеют интенсивность пикселей, превышающую пороговое значение.Активированные клетки показали явное увеличение интенсивности флуоресценции (рисунок B в файле S1), а измеренное время активации не было чувствительным к небольшим изменениям пороговой интенсивности, используемой при анализе изображений, см. Рисунок C в файле S1.
Как упоминалось ранее [9], существует два основных типа механизмов перекрестных помех AHL между видами микробов, как показано на рис. 1D. В контексте перекрестных помех отправители будут создавать родственный AHL для получателей, в то время как взаимодействующие будут производить не родственный вариант сигнала, который связывается с рецептором LuxR.Для возбуждающих перекрестных помех взаимодействующие виды стимулируют активацию приемников, в то время как для ингибирующих перекрестных помех взаимодействующие виды подавляют активацию.
Введение в систему «отправитель-получатель» интерактора должно сдвинуть время активации получателей. В качестве начального положительного контроля, показанного на фиг. 2, клетки-отправители, содержащие luxI , были добавлены на лужайку приемников, чтобы проверить уменьшение времени активации на 0,75 часа для штамма взаимодействия, производящего родственный сигнал.В отрицательном контроле, где интеракторный штамм представляет собой дикий тип E . coli штамм-хозяин, не продуцирующий сигнал AHL, время активации не изменилось. Этот результат указывает на то, что пространство, занимаемое взаимодействующими ячейками, не повлияло на реакцию принимающих ячеек (см. Рисунок D в файле S1).
Рис. 2. Анализ фиксирует диапазон перекрестных помех.
Схема анализа перекрестных помех. Кимограмма экспрессии флуоресценции в клетках-приемниках в положении 2 мм от колонии-отправителя для пяти различных условий.Квадратики показывают флуоресцентные изображения, снятые между 4 и 6 часами. Две верхние строки демонстрируют, что добавление штамма интерактора, который не производит никакого сигнала, WT, не изменило время активации. Добавление интеракторного штамма, содержащего ген сигнальной синтазы (три нижних условия), сдвигало время активации.
https://doi.org/10.1371/journal.pcbi.1005809.g002
Мы протестировали перекрестные помехи от неродственных сигналов, представив E .Штамм coli , содержащий гены синтазы rhlI или lasI , продуцирующий C4-HSL и 3-оксо-C12-HSL в качестве своих основных продуктов, соответственно [9,23]. Мы выбрали эти две цепи AHL, основываясь на доказательствах как возбуждающих, так и тормозных перекрестных помех с системой LuxI / R из предыдущих исследований [17,20,23,29]. Как показано на рис.2, когда модель E . coli LasI штамм был введен как штамм взаимодействия, приемники активировали RFP раньше по сравнению с контролем без перекрестных помех.Когда модель E . coli RhlI штамм был введен как штамм-взаимодействующий, приемники активировали RFP позже. Эти первоначальные тесты подтвердили, что в нашем анализе могут наблюдаться как тормозящие, так и возбуждающие перекрестные помехи. Как видно на рисунке E в файле S1, введение этих штаммов-интеракторов не влияет на рост штаммов-отправителей или получателей, подтверждая вывод о том, что наблюдаемый эффект был вызван перекрестными помехами между AHL и рецептором.
Измерение масштабирования задержки перекрестных помех с составом сообщества
Задержка активации штамма-приемника в зависимости от количества взаимодействующих клеток измерялась путем изменения количества штамма-приемника, загруженного на планшет.Количество взаимодействующего штамма, добавленного в планшет, фиксируется как отношение взаимодействующего элемента к принимающему, которое определяется как отношение количества взаимодействующих клеток, загруженных в анализ на планшете, к количеству принимающих клеток, загруженных в тестовый планшет. Количество приемников всегда оставалось постоянным и составляло 10 8 ячеек. Как показано на фиг. 3, сдвиг времени активации был пропорционален количеству взаимодействующих клеток. Кривые активации показаны для случаев возбуждающего перекрестного взаимодействия (рис. 3A), с LasI в качестве штамма взаимодействия, и ингибирующего перекрестного взаимодействия (рис. 3B), с RhlI как штамм-взаимодействующий элемент.Эти эксперименты позволяют количественно оценить, как распространение фронта активации зависит от состава сообщества, как с точки зрения типов производимых сигналов, так и относительного количества каждого напряжения взаимодействия в окружающей среде.
Рис. 3. Зависимость динамики активации от количества межклеточных клеток.
A. Добавление штамма LasI в качестве штамма-взаимодействия сокращает время активации по сравнению со случаем отсутствия перекрестных помех. B. Добавление штамма RhlI в качестве интерактора увеличивает время активации по сравнению со случаем отсутствия перекрестных помех.В обоих случаях сдвиг во времени активации был пропорционален количеству добавленной деформации интерактора, как определено соотношением интерактора к получателю (подробности см. В тексте). Экспериментальные данные взяты из трех независимых измерений. Графики на заднем плане показывают тенденцию изменения данных и предназначены исключительно для ориентации глаз.
https://doi.org/10.1371/journal.pcbi.1005809.g003
Моделирование перекрестных помех, обнаруживаемых кворумом, с помощью уравнения реакции-диффузии
В этом разделе мы построим математическую модель для изучения корреляций связывания сигнала на микроуровне с рецептором и пространственно-временных паттернов экспрессии генов на макроуровне в системе, включающей перекрестные помехи, воспринимающие кворум.В предыдущей работе, выполненной в [21,22,28,30–32], авторы реализовали уравнение логистического роста (1) и модель реакции-диффузии (2) для моделирования продукции сигнала от растущих клеток. Уравнение логистического роста учитывает переходное поведение плотности клеток n i , которая растет со скоростью μ на клетку. Поскольку носитель имеет ограниченное количество ресурсов, общая плотность ячеек (отправители + получатели + взаимодействующие), n T , приблизится к плотности насыщенных ячеек s.
(1)
Первоначально отправители производят AHL с базальной скоростью ? b на ячейку. Передающие AHL с концентрацией c s , диффундируют от ячеек с коэффициентом диффузии D c и разлагаются средой со скоростью d а . ? объясняет увеличение продукции АГЛ при наличии сигнала. Для отправителей активность (A) определяет, как эта активированная скорость производства сигнала из-за изменений в продукции синтазного белка зависит от концентраций нескольких AHL.
(2)
В присутствии взаимодействующих AHL, транскрипционная активность будет модулироваться из-за связывания AHL с рецепторами LuxR. Каждый сигнал по-разному влияет на активность, как в способности связываться с рецептором, так и в последующем влиянии такого связывания на экспрессию генов, контролируемых кворумом. Способность AHL связываться с рецептором LuxR зависит от энергии связывания и локальной концентрации каждой AHL. При моделировании мы рассмотрели вероятность связывания AHL с рецептором и ввели вес, чтобы учесть последующее влияние каждого варианта AHL на экспрессию гена.Таким образом, деятельность принимает форму:
(3)
где g — количество рецепторов на клетку, i — индекс, описывающий тип AHL, c i — концентрация i th AHL, P ( c i ) — вероятность связывания AHL с рецептором, w i — весовой параметр, j — общее количество взаимодействующих AHL. Вероятность связывания объясняет различия в аффинности связывания каждого варианта сигнала с рецептором, а также конкуренцию за связывание нескольких сигналов с одним и тем же рецептором.Количество рецепторов (g) изменяется от базового уровня от 100 до 600, поскольку уровни экспрессии генов увеличиваются из-за накопления сигнала. Чтобы смоделировать этот плавный переход, мы использовали функцию Хилла, см. Таблицу A в файле S1. Нулевой или положительный уровень транскрипционной активности является только физическим, поэтому веса также должны быть больше или равны нулю.
Весовой параметр ( w i ) связывает количество комплексов AHL-рецептор со степенью регуляции гена, при этом большие положительные веса указывают на то, что комплексы, образованные этими AHL, приводят к сильной активации регулируемых генов, воспринимающих кворум, в то время как веса, близкие к нулю, приводят к ингибированию этих генов.Вес определяется сродством связанного рецептора к промоторной области регулируемых генов, воспринимающих кворум, эффективностью транскрипции и трансляции регулируемых генов, воспринимающих кворум, и скоростью диссоциации комплекса AHL-рецептор. Детерминированный весовой подход Больцмана был применен для расчета вероятностей связывания рецепторов на основе концентраций AHL и энергий связывания рецепторов [33], см. Математическую модель в тексте A в файле S1 и на рисунках F-H в файле S1.Значения параметров, приведенные в таблице A в файле S1, были измерены в контрольных экспериментах или получены из предыдущих экспериментальных исследований [19,31,34–36].
Поскольку интеракторный штамм не продуцировал никаких рецепторов, продукция сигнала была конститутивной и не включала положительную обратную связь от уровня AHL. Активность (A) для интеракторов была равна нулю, и производство сигнала происходило на базальном уровне,
(4)
Предполагалось, что основная скорость производства равна максимальной производительности отправителей.Эти уравнения решались методом конечных разностей. Прогнозы модели были получены с учетом переходного поведения концентраций АГЛ. При моделировании мы рассматривали две концентрические окружности; внутренний круг имеет радиус 1 мм, а внешний круг — 10 мм. Плотность ячеек регулируется уравнением (1), динамика сигналов отправителей регулируется уравнениями (2) и (3), а взаимодействующие элементы — уравнением (4). Начальные условия для моделирования были выбраны в соответствии с экспериментальными условиями.Как и в экспериментах, изначально во внутренний круг было добавлено 10 7 клеток-отправителей. Предполагалось, что внутренний круг имеет начальную концентрацию АГЛ 70 нМ. Внешний круг имеет переменную смесь приемников и взаимодействующих устройств, равномерно распределенных в пространстве. Во всех случаях было 10 8 ячеек-приемников. Начальная концентрация интеракторной АГЛ во внешнем круге составляла 70 нМ [31]. При моделировании величина деформации взаимодействующего элемента была скорректирована в соответствии с соотношением взаимодействующего элемента к приемному устройству.
На основе диффузных градиентов сигналов, созданных отправителями и взаимодействующими видами, активность приемников была рассчитана с использованием,
(5)
где P ( c S ) — это вероятность того, что AHL от отправителя будут связываться с рецептором, P ( c int ) — это вероятность того, что AHL из интерактор будет связываться с рецептором, w s — это вес, связанный с AHL отправителя, и w int — вес, связанный с AHL интерактора.Активность приемников модулирует продукцию флуоресцентного репортера гена (RFP), поскольку репортерный ген транскрибируется промотором, регулируемым сигнальным рецептором.
Уровень активности приемников действует как индикатор изменений в экспрессии генов в результате перекрестных помех. Поэтому мы определяем пороговый уровень активности для активации экспрессии гена и использовали уравнение (5), чтобы отслеживать, превышает ли уровень активности приемников этот порог. При моделировании пороговая активность принималась равной половине максимального уровня активности при отсутствии перекрестных помех.
Устойчивость передачи сигнала к перекрестным помехам
Для получения весов сигналов передающего AHL и двух взаимодействующих AHL результаты моделирования были подогнаны с использованием экспериментальных данных с рис. 3. Весовой параметр (w 1 ) для сигнала связывания 3-оксо-C6-HSL с LuxR рецептор был подобран с использованием экспериментальных данных для отсутствия перекрестных помех. Для этой цели использовался нелинейный метод наименьших квадратов, см. Рисунок Ia в файле S1. Дополнительные весовые коэффициенты необходимы для учета каждого взаимодействующего AHL.Чтобы идентифицировать весовые параметры взаимодействующих AHL, экспериментальные графики, показанные на рис. 3, были подогнаны для случая отношения взаимодействующего элемента к приемнику 0,9 с использованием нелинейной аппроксимации методом наименьших квадратов (рис. I в файле S1). Рассчитанные веса для сигналов, производимых деформациями взаимодействующего элемента LasI и RhlI, показаны в таблице 1. С использованием этих весов были смоделированы кривые активации для отношения взаимодействующего элемента к приемнику 0,2 и 0,5, как показано на рисунках 4A и 4B, что дает масштабирование, аналогичное приведенному на рис. экспериментальные данные представлены на рис.3.
Рис. 4. Устойчивость активации экспрессии гена к перекрестным помехам.
А, Б. Теоретические прогнозы реакции принимающих клеток на возбуждающие или тормозящие перекрестные помехи. Экспериментальные данные на рис. 3 показаны для сравнения. C. Сравнение экспериментальных измерений активации экспрессии гена в тесте на планшете с прогнозами, сделанными с использованием модели реакции диффузии. Линии показывают прогнозируемое изменение времени активации на нескольких расстояниях от колонии-отправителя в зависимости от количества штамма взаимодействия, добавленного в планшет.Прогнозы были сделаны с использованием экспериментально рассчитанных весов перекрестных помех для напряжений взаимодействия RhlI и LasI. Точки данных показывают экспериментальные измерения на выбранных расстояниях из рис. 3.
https://doi.org/10.1371/journal.pcbi.1005809.g004
Проверенная модель перекрестных помех позволила исследовать устойчивость распространения сигнала. Распространение сигнала в присутствии переменных уровней перекрестных помех моделировалось как для штаммов взаимодействия LasI, так и для RhlI.На рис. 4C показана прогнозируемая задержка активации деформации приемника на расстояниях 0, 2, 5, 7 и 10 мм для отношения взаимодействующего элемента к значениям приемника между 0 и 1. Точки данных показывают экспериментальные измерения активации ответа RFP. на таких расстояниях и уровнях перекрестных помех, что свидетельствует о хорошем согласии с прогнозами модели.
Используя модель, мы спрогнозировали чувствительность динамики сигналов к изменениям параметров модели, включая скорость роста клеток (рисунок J в файле S1), скорость производства сигнала (рисунок K в файле S1), коэффициент диффузии (рисунок L в файле S1). , а также скорость ухудшения сигнала (рис. M в файле S1).Время активации сильно зависит от коэффициента диффузии, скорости образования сигнала и скорости деградации сигнала, поскольку вместе эти параметры задают профиль концентрации сигнала. Скорость роста не влияла на время активации, вероятно, из-за того, что клетки загружали на планшет с плотностью, близкой к насыщению, поэтому во время эксперимента имело место небольшое количество делений. Кроме того, мы наблюдаем, что перекрестные помехи сильно коррелируют с энергией связи и параметром веса, см. Рисунок N и рисунок O в файле S1.Активность штамма-приемника для переменных концентраций сигналов, создаваемых штаммом-отправителем и штаммом интерктора, также представлена на рисунке P в файле S1. Модель предсказывает, что на активность отправителей не влияет обмен сигналами с взаимодействующими устройствами, см. Рисунок Q в файле S1.
Измерение перекрестного потенциала диких изолятов
Анализ также позволяет измерять перекрестные помехи с дикими изолятами. В качестве первоначального теста были измерены перекрестные помехи с Pseudomonas aeruginosa дикого типа, как показано на фиг. 5A.Присутствие Pseudomonas aeruginosa при соотношении интерактора и приемника 0,9 задерживает активацию на несколько часов. Поскольку гены las и rhl являются производными от Pseudomonas aeruginosa , весовые параметры извлечены из E . coli, штаммы интерактора были использованы для прогнозирования ожидаемой задержки активации в результате перекрестных помех с этими двумя системами. Мы моделировали взаимодействия с гипотетическим штаммом взаимодействия, содержащим как las , так и rhl .Здесь активность приемников будет,
(6)
Рис. 5. Тестирование модели и эксперименты с естественными изолятами.
A. Сравнение результатов моделирования с экспериментальными результатами с Pseudomonas aeruginosa в качестве штамма взаимодействия. Данные из штаммов взаимодействия LasI и RhlI были использованы для прогнозирования комбинированного влияния цепей обнаружения кворума LasI и RhlI в P . aeruginosa . Линия влияния роста LasI + RhlI + добавляет экспериментально измеренное снижение скорости роста E . coli в наличии P . aeruginosa к модели, см. Рисунки R-T в файле S1. B. Планшетный анализ использовали для измерения интерференционного потенциала четырех диких бактериальных изолятов при соотношении интерактора к получателю 0,9. Линии, показанные для направления взгляда, экспоненциально соответствуют данным.
https://doi.org/10.1371/journal.pcbi.1005809.g005
На рис. 5A мы заметили, что тенденция моделируемой кривой активации аналогична экспериментальным результатам, показывая отложенную активацию и более пологую кривую активации по всему периметру. пластина.Прогнозируемая задержка была короче экспериментально измеренной примерно на 2 часа. В файле S1 выполняется дополнительное моделирование, чтобы определить, есть ли задержка в росте штамма-отправителя или влияние штамма-отправителя AHL на продукцию AHL в P . aeruginosa может вносить вклад в дополнительную задержку активации. Снижение скорости роста штамма-отправителя при совместном культивировании с P . aeruginosa , было подтверждено измерениями роста, см. Рисунок R в файле S1.На рис. 5А показан прогноз модели, учитывающий влияние роста. Хотя активация QS была отложена, одних только ростовых взаимодействий было недостаточно для воспроизведения 2-часовой задержки активации, см. Текст B, рисунок S и рисунок T в файле S1.
Затем был протестирован потенциал перекрестных помех четырех дополнительных диких изолятов. Эти четыре препарата были добавлены в планшет для анализа при соотношении взаимодействующего вещества и приемника 0,9. Секвенирование 16S рРНК идентифицировало виды дикого типа как Aeromonas hydrophila , Aeromoans veronii , Pantoea agglomerans и Pantoea vagans .О способности этих штаммов продуцировать АГЛ сообщалось ранее [37–42]. На рис. 5В мы наблюдали, что, когда A. veronii или P. vagans были добавлены к лужайке клеток-приемников, активация RFP в приемниках происходила раньше, по сравнению с отсутствием штамма взаимодействия. A. hydrophila и P. agglomerans задерживают активацию. Степень перекрестных помех была различной для каждого вида, предполагая, что на активацию генетической экспрессии в различных сообществах, вероятно, влияют перекрестные помехи различной силы от нескольких видов.
Обсуждение
Наши результаты дают новое понимание передачи сигналов в смешанных сообществах бактерий. Адаптировав подход, использованный в предыдущих исследованиях [24,25,28,43–45], мы создали анализ планшета типа отправитель-получатель для количественной оценки активации экспрессии генов из-за AHL-опосредованной передачи сигналов в присутствии нескольких штаммов, продуцирующих сигнал. В ходе анализа измерялась устойчивость конкретных сигнальных сетей к помехам со стороны штамма, продуцирующего чужеродные сигнальные молекулы.При сравнении анализа, основанного на пространственной реакции-диффузии, с хорошо перемешанными системами, мы обнаружили, что пространственный анализ способен дифференцировать штамм-взаимодействующий элемент, который производит неродственный сигнал, от штамма, который вырабатывает фермент, разрушающий сигнал, см. Рисунок U в S1. Файл. Хотя ранее сообщалось о перекрестных помехах между сетями восприятия кворума [9,11,12,23,46,47], наше титрование взаимодействующего штамма выявило чувствительность опосредованной сигналом экспрессии генов в пространственно распределенной сети к помехам.Как показано на рис. 4C, на расстояниях 2 мм или меньше кратное изменение составляет менее 10% даже для коэффициентов перекрестных помех, равных 1. На расстояниях 10 мм время активации изменилось примерно на 10% при только 20% соотношении взаимодействующих элементов. к получателю. Эти числа предполагают, что генетическая активность, основанная на кворуме, в значительной степени устойчива к помехам и что любой многочисленный вид должен активировать свою сеть распознавания кворума в типичной системе. Наши результаты относятся к вмешательству LasI и RhlI в систему LuxRI QS, и пока неясно, развились ли некоторые системы AHL с разными уровнями устойчивости или чувствительности к определенным неродственным сигналам.
Модель демонстрирует устойчивость сети AHL к помехам отчасти из-за различий в энергиях связывания родственных и не родственных сигналов. Рисунок N в файле S1 показывает, что по мере ослабления энергии связывания неродственного сигнала перекрестные помехи от неродственного сигнала мало влияют на регуляцию гена. Отношение взаимодействующих к принимающим клеткам также влияет на надежность. Как показано на рисунке V в файле S1, когда количество взаимодействующих клеток значительно превышает количество принимающих клеток, устойчивость теряется, и экспрессия регулируемых LuxR / I генов задерживается на несколько часов.Третий фактор, влияющий на надежность, — это вес взаимодействия для неродственного сигнала, как показано на рисунке O в файле S1. Дальнейшая работа должна дополнительно охарактеризовать диапазон весов взаимодействия, присутствующих в реальных системах. Лучшее понимание устойчивости обмена сигналами в смешанных популяциях было бы полезным для реализации генных цепей, воспринимающих кворум, в синтетических микробных сообществах [2,15,18–20,44].
Наши экспериментальные измерения помогли в разработке детальной модели для предсказания основанной на AHL динамики передачи сигналов в смешанных популяциях.Модель учитывает перекрестные помехи между штаммами, используя один параметр, называемый весом, который мы вычисляем из экспериментов для данного рецептора для каждой комбинации сигналов. Этот вес учитывает нижестоящие регуляторные последствия связывания рецептора с AHL и может быть связан с фундаментальными процессами, такими как димеризация рецептора, взаимодействия между рецептором, ДНК и РНК-полимеразой, а также транскрипция и трансляция регулируемых AHL генов. . Мы обнаружили, что модель, использующая одно значение веса, хорошо согласуется с экспериментальной динамикой активации, охватывающей более 1 см пространства с переменным количеством помех.Это близкое соответствие между моделью и экспериментами предполагает, что модель может быть реализована для изучения перекрестных помех, воспринимаемых кворумом, в более сложных и реалистичных контекстах, например, при наличии более двух штаммов, когда клетки неоднородно распределены в пространстве или даже при транспортировке. динамика пространственно зависима [21,25,30]. Поскольку эксперименты в этих условиях были бы сложными, наш анализ и модель обеспечивают прямой путь к предсказанию динамики передачи сигналов в сложных условиях.В дополнение к прогнозированию динамики в сложных условиях, как упоминалось выше, модель позволила исследовать, как устойчивость к помехам может появиться путем настройки параметров, которые регулируют реакцию на обмен сигналами. Устойчивость может быть достигнута, если рецептор эволюционировал, чтобы связывать неродственный сигнал гораздо слабее, чем родственный сигнал. Разница в энергиях связывания рецептора между неконфекционными и родственными сигналами, необходимыми для устойчивости, зависит от количества межклеточных клеток и влияния неродственного сигнала на экспрессию гена, что отражено в весовом термине.Некоторые рецепторы, возможно, развили достаточную степень несоответствия связывания на основе взаимодействующих штаммов и обычно встречающихся неродственных сигналов.
Наш анализ интерференции сигналов с дикими видами выявил широкий спектр перекрестных помех в естественных популяциях. Мы обнаружили как возбуждающие, так и тормозящие перекрестные помехи внутри наших изолятов, а также различную степень перекрестных помех с системой распознавания кворума luxRI. Предыдущие измерения также показали, что непородные AHL могут взаимодействовать с рецепторами, такими как LuxR, в различной степени [9,23].Поскольку мы используем дикий изолят непосредственно вместо очищенного сигнального, бесклеточного супернатанта или синтетических штаммов-продуцентов, мы фиксируем как прямые, так и косвенные сигнальные взаимодействия с взаимодействующим штаммом. Примеры непрямых взаимодействий включают модуляцию скорости роста и пути регуляции генов, а также эволюционирующие пространственные распределения интерактора. Мы попытались в случае взаимодействия с Pseudomonas aeruginosa указать источник этих косвенных сигнальных взаимодействий, независимо учитывая влияние каждого сигнала AHL, производимого штаммом интерактора, и влияния роста штамма интерактора на принимающие клетки.Хотя модель предсказывала увеличенную задержку активации из-за эффектов роста, как показано на рис. 5A, все еще существуют дополнительные неизвестные в настоящее время взаимодействия, которые еще больше задерживают активацию. Мы предполагаем, что рецептор QscR, находящийся в Pseudomonas aeruginosa , может поглощать AHL-отправители и вносить вклад в эту задержку [48], хотя другие регуляторные взаимодействия между видами, не основанные на AHL, вероятно, вносят вклад в динамику передачи сигналов. В будущих усилиях следует попытаться разделить прямые и косвенные взаимодействия, которые влияют на передачу сигналов, чтобы улучшить нашу способность предсказывать динамику передачи сигналов в реальных популяциях.Кроме того, для некоторых экологических ниш динамика роста и движения клеток могут влиять на градиенты AHL неожиданным образом, и эти факторы должны быть включены в любую будущую работу, чтобы понять динамику передачи сигналов в сложных средах [49,50]. Использование анализа для широкого выборочного анализа взаимодействий между известной системой рецепторов AHL, такой как luxRI, и продуцентами диких сигналов, должно дать новое понимание паттернов перекрестных помех в реальных условиях и их последствий для регуляции кворума на уровне экосистемы.
Материалы и методы
Бактериальные штаммы и плазмиды
В таблице 2 мы представили подробную информацию о штаммах бактерий, использованных в этом исследовании. Штамм хозяина, используемый для отправителя, получателя и взаимодействующих, — это Escherichia coli NEB 5-alpha. Основными сигналами QS являются 3-оксо-C6 HSL для штамма-отправителя, 3-оксо-C12 HSL для штамма взаимодействия LasI и C4-HSL для штамма взаимодействия RhlI, см. Рисунок A в файле S1 для получения дополнительных сведений. Плазмиды были либо получены от Addgene [21], либо сконструированы с использованием сборки Гибсона (New England Biolabs).
Условия культивирования
Штаммы бактерий инокулировали из замороженных исходных материалов в пробирке Falcon на 12 мл с 5 мл бульона LB с соответствующими антибиотиками. Посевной материал выращивали на шейкере при 220 об / мин при 37 ° C в течение 16 часов. Клетки ресуспендировали в свежей среде для удаления сигнала в супернатанте. Использовались культуры поздней логарифмической фазы, так что определение кворума штамма-отправителя активировалось перед измерением в анализе на планшете.
Планшетный анализ
Планшетный анализ был настроен, как описано в Silva et al [35].Взаимодействующий штамм смешивали со 100 мкл приемников в центрифужной пробирке на 1,5 мл и наносили на верхнюю часть пластин с агаром 2,5% LB, используя стерильные стеклянные шарики 4 мм. Рисунок W в файле S1 показывает, что пространственное распределение клеток оставалось смешанным во время анализа.
Измерения под микроскопом
Для получения изображений использовался флуоресцентный микроскоп Nikon eclipse TI. Эксперименты проводились при 37 ° C с использованием камеры с регулируемой температурой. Образцы были сфотографированы при 20-кратном увеличении.Для регистрации активации RFP в ячейках-получателях изображения RFP снимались каждые 15 минут в течение 16 часов на 30 различных расстояниях от колонии-отправителя. Время активации было рассчитано для каждой позиции. Время экспозиции составляло 1 с для RFP и 500 мс для GFP. Существенного фотообесцвечивания не наблюдалось.
Каждое сделанное изображение было сохранено в формате .tiff и проанализировано с использованием специального кода Matlab. К изображениям RFP был применен низкий порог, чтобы идентифицировать расположение ячеек приемника в каждом изображении.Верхний порог использовался для идентификации получателей, которые активировали RFP. Для каждой временной точки и положения вычислялась доля пикселей ячейки выше порога активации RFP. Если доля активированных пикселей превышала 10%, эта позиция включалась как часть активированной области, см. Рисунок C в файле S1.
Измерения роста
Для получения кривых роста ночные культуры разводили от 1 до 1000 в среде LB и проводили селективное посевное покрытие для измерения плотности клеток с течением времени.Чтобы получить кривые роста из смесей штаммов, каждый штамм имел уникальный маркер устойчивости и был помещен на соответствующий планшет для отбора.
Измерения планшет-ридера
Планшетный ридер
Tecan Infinite m200 Pro использовали для измерения скорости роста и активации флуоресценции в хорошо перемешанных условиях. Клетки выращивали до поздней логарифмической фазы, разводили в 1000 раз чистой средой LB и культивировали в течение дополнительных 3 часов. После трех часов роста 200 мкл этих клеток с ранней логарифмической фазой загружали в 96-луночный планшет с плоским дном.Планшет вставляли в устройство для чтения планшетов, установленное на 37 ° C, и измеряли оптическую плотность и интенсивность флуоресценции каждые 15 минут в течение 16 часов. Измерения оптической плотности проводились на длине волны 600 нм. Для измерений GFP использовали длину волны 485 нм для возбуждения и длину волны 515 нм для излучения. Для измерений флуоресценции RFP использовали длину волны 590 нм для возбуждения и длину волны 650 нм для излучения.
Благодарности
Мы благодарим Шона Лима и Сяокан Го за помощь в выделении и идентификации штаммов.
Ссылки
- 1.
Миллер М.Б., Басслер Б.Л. Анализ кворума бактерий. Annu Rev Microbiol. 2001; 55: 165–99. pmid: 11544353 - 2.
Резерфорд С.Т., Басслер Б.Л. Зондирование бактериального кворума: его роль в вирулентности и возможности контроля. Cold Spring Harb Perspect Med. 2016; 1–26. - 3.
Нилсон К. Х., Гастингс Дж. У. Исследование кворума в глобальном масштабе: огромное количество биолюминесцентных бактерий образуют молочные моря. Appl Environ Microbiol.2006. 72: 2295–2297. pmid: 16597922 - 4.
Боедикер Дж., Нилсон К. Микробная коммуникация через зондирование кворума. IEEE Trans Mol Biol Multi-Scale Commun. 2016; 1: 310–320. - 5.
Тереза Р., Иглевски Б.Н. Определение бактериального кворума в патогенных отношениях. Infect Immun. 2000; 68: 4839–4849. pmid: 10948095 - 6.
Уильямс П., Винзер К., Чан В. К., Камара М. Посмотрите, кто говорит: общение и определение кворума в мире бактерий. Philos Trans R Soc London B Biol Sci.2007; 362: 1119–1134. pmid: 17360280 - 7.
Раджпут А., Каур К., Кумар М. SigMol: Репертуар сигнальных молекул, воспринимающих кворум в прокариотах. Nucleic Acids Res. 2016; 44: D634 – D639. pmid: 264 - 8.
Винендал Э., Бронселер А., Ниландт Дж., Д’Хондт М., Сталманс С., Брак Н. и др. База данных Quorumpeps: химическое пространство, микробное происхождение и функциональность пептидов, воспринимающих кворум. Nucleic Acids Res. 2013; 41: 655–659. pmid: 23180797 - 9.
Рэмпиони Дж., Леони Л., Уильямс П.Биоорганическая химия Искусство антибактериальной войны: обман через вмешательство в коммуникацию, опосредованную кворумом. Bioorg Chem. Elsevier Inc .; 2014; 55: 60–68. pmid: 24823895 - 10.
Хьюз Д. Т., Сперандио В. Передача сигналов между царствами: связь между бактериями и их хозяевами. Nat Rev Microbiol. 2008. 6: 111–120. pmid: 18197168 - 11.
МакКлин К.Х., Винсон М.К., Фиш Л., Тейлор А., Чхабра С.Р., Камара М. и др. Определение кворума и Chromobacterium violaceum: использование производства и ингибирования виолаацеина для обнаружения лактонов N-ацилгомосерина.Микробиология. 1997; 143 (Pt 1: 3703–11. Pmid: 9421896 - 12.
Перес П.Д., Вайс Дж. Т., Хаген С. Дж. Шум и перекрестные помехи в двух входах Vibrio fischeri, распознающих кворум. BMC Syst Biol. 2011; 5: 153. pmid: 21959018 - 13.
Holden MTG, Chhabra SR, De R, Stead P, Bainton NJ, Hill PJ и др. Перекрестный разговор на основе определения кворума: выделение и химическая характеристика циклических дипептидов из синегнойной палочки и других грамотрицательных бактерий. Mol Microbiol. 1999; 33: 1254–1266.pmid: 10510239 - 14.
Грант П.К., Далчау Н., Браун Дж. Р., Федеричи Ф., Радж Т. Дж., Йорданов Б. и др. Ортогональная межклеточная передача сигналов для запрограммированного пространственного поведения. Mol Syst Biol. 2016; 12: 849: 1–13. - 15.
Гао Й, Сун Дж., Ху Б., Чжан Л., Лю Ку, Лю Ф. Ген luxS участвует в производстве AI-2, патогенности и некоторых фенотипах у Erwinia amylovora. Curr Microbiol. 2009; 58. pmid: 18820970 - 16.
Lasarre B, Federle MJ. Использование зондирования кворума для запутывания бактериальных патогенов.Microbiol Mol Biol Rev.2013; 77: 73–111. pmid: 23471618 - 17.
Дэвис Р.М., Мюллер Р.Ю., Хейнс К.А., Дэвис Р.М., Фултон И.А. Может ли естественное разнообразие кворумов продвинуть вперед синтетическую биологию? Фронт Bioeng Biotechnol. 2015; 3: 1–10. - 18.
Скотт С.Р., Хэсти Дж. Коммуникационные модули для анализа кворумов для микробных консорциумов. ACS Synth Biol. 2016; 5: 969–977. pmid: 27172092 - 19.
Teng S-W, Schaffer JN, Tu KC, Mehta P, Lu W, Ong NP и др. Активная регуляция соотношения рецепторов контролирует интеграцию сигналов, воспринимающих кворум, у Vibrio harveyi.Mol Syst Biol. Издательская группа «Природа»; 2011; 7: 491. pmid: 21613980 - 20.
Кантон Б., Лабно А., Энди Д. Уточнение и стандартизация синтетических биологических частей и устройств. Nat Biotechnol. 2008; 26: 787–793. pmid: 18612302 - 21.
Данино Т., Мондрагон-Паломино О., Цимринг Л., Хэсти Дж. Синхронизированный кворум генетических часов. Природа. Издательская группа «Природа»; 2010. 463: 326–330. pmid: 200 - 22.
Приндл А., Самайоа П., Разинков И., Данино Т., Цимринг Л.С., Хэсти Дж.Чувствительный массив радикально связанных генетических биопикселей. Природа. 2012; 481: 39–44. - 23.
Ву Ф., Менн Д. Д., Ван Х. Чувствительные к кворуму синтетические схемы, управляемые перекрестными помехами: от унимодальности к тримодальности. Chem Biol. Elsevier Ltd; 2014; 21: 1629–1638. pmid: 25455858 - 24.
Диланджи Г.Е., Лангебрейк Дж., Линхир П. Де, Стивен Дж. Активация кворума на расстоянии: пространственно-временные паттерны регуляции генов от диффузии сигнала аутоиндуктора. J Am Chem Soc. 2012; 1–17. - 25.
Basu S, Gerchman Y, Collins CH. Синтетическая многоклеточная система для запрограммированного формирования паттернов. Природа. 2005; 434: 1130–1134. pmid: 15858574 - 26.
Kong W, Celik V, Liao C, Hua Q, Lu T. Программирование группового поведения бактериальных сообществ с помощью синтетической клеточной коммуникации. Биоресурсы Биопроцесс. 2014; 1: 24. - 27.
Рамальо Т., Мейер А., Мюкл А., Капснер К. Анализ отдельных клеток бактериальной системы отправителя-получателя.PLoS One. 2016; 11: 1. pmid: 26808777 - 28.
Тровато А., Сено Ф., Занардо М., Альбергини С., Тонделло А., Скуартини А. Кворум против диффузного зондирования: количественный анализ релевантности поглощающих или отражающих границ. FEMS Microbiol Lett. 2014; 352: 198–203. pmid: 24484313 - 29.
Сонг Х., Одзаки Дж., Коллинз С.Х., Барнет М., Арнольд Ф.Х., Quake SR. Синтетическая экосистема хищник – жертва Escherichia coli. Mol Syst Biol. 2008; 1–8. pmid: 18414488 - 30.Langebrake JB, Dilanji GE, Hagen SJ, Leenheer P De. Бегущие волны в ответ на распространяющийся сигнал зондирования кворума в пространственно протяженных бактериальных колониях. J Theor Biol. 2014; 363: 53–61. pmid: 25109591
- 31.
Фекете А., Каттлер С., Ротбаллер М., Хенсе Б.А., Фишер Д., Буддрус-Шиман К. и др. Динамическое регулирование продукции и деградации N-ацил-гомосерин-лактона в Pseudomonas putida IsoF. FEMS Microbiol Ecol. 2010; 72. pmid: 20100181 - 32.
Пай А, Танучи Й, Ю Л.Оптимальность и надежность контроля кворума (QS) — опосредованная регуляция дорогостоящего фермента общественного блага. Proc Natl Acad Sci. 2012; 109. pmid: 23144221 - 33.
Ван Вален Д., Хатая М., Филлипс Р. Биохимия на поводке: роль длины и геометрии привязи в белках интеграции сигналов. Biophys J. Биофизическое общество; 2009; 96: 1275–1292. pmid: 19217847 - 34.
Зенглер К., Палссон Б.О. Дорожная карта развития сообщества системной (уютной) биологии. Nat Rev Microbiol.Издательская группа «Природа»; 2012; 10: 366–372. pmid: 22450377 - 35.
Сильва К.П., Челламуту П., Бёдикер Дж. К. Уничтожение сигнала настраивает зону активации в пространственно распределенных сигнальных сетях. Biophys J. Биофизическое общество; 2017; 112: 1037–1044. pmid: 28297640 - 36.
Гнанендра С., Анусуя С., Натараджан Дж. Молекулярное моделирование и анализ активного сайта гомолога SdiA, предполагаемого кворума сенсора патогенности сальмонеллы typhimurium, выявляют специфические паттерны связывания регуляторов транскрипции AHL.Модель J Mol. 2012; 18: 4709–4719. pmid: 22660944 - 37.
Гард С., Бьярнсхольт Т., Гивсков М., Якобсен Т.Х., Хентцер М., Клауссен А. и др. Регламент определения кворума у Aeromonas hydrophila. J Mol Biol. Elsevier Ltd; 2010; 396: 849–857. pmid: 20064524 - 38.
Свифт С., Карлышев А.В., Фиш Л., Дюрант Е.Л., Винсон М.К., Чхабра SRAM и др. Определение кворума у Aeromonas hydrophila и Aeromonas salmonicida: идентификация гомологов LuxRI AhyRI и AsaRI и их когнитивных сигнальных молекул N-ацилгомосерин-лактона.J Bacteriol. 1997; 179: 5271–5281. pmid: 9286976 - 39.
Джангид К., Парамешваран П.С., Шоуч Ю.С., Коллекция MC. Вариант системы зондирования кворума в Aeromonas veronii MTCC 3249. Датчики. 2012; 12: 3814–3830. pmid: 22666003 - 40.
Чан X, Как К., Инь В., Чан К. Опосредованное N-ацил гомосерин-лактоном кворум-зондирование в штамме Sobria биовара Aeromonas veronii 159: Идентификация гомологов LuxRI. Front Cell Infect Microbiol. 2016; 6: 1–6. - 41.
Хосни Т., Моретти С., Девескови Г., Суарес-Морено З. Р., Фатми М.Б., Гуарначча С. и др.Совместное использование сигналов распознавания кворума и роль межвидовых сообществ в бактериальном заболевании растений. ISME J. 2011; 5: 1857–1870. pmid: 21677694 - 42.
Chalupowicz L, Manulis-Sasson S, Itkin M, Sacher A, Sessa G, Barash I. Система определения кворума влияет на развитие желчи, вызванное Pantoea agglomerans pv. гипсофилы. Мол Растительный Микроб Взаимодействовать. 2008; 21: 1094–1105. pmid: 18616406 - 43.
Писарро Г., Гриффит Д., Ногера Д. Количественная модель клеточного автомата для биопленок.J Environ Eng. 2001; 127. - 44.
Конг Вентао, Бланшар Эндрю Э., Чэнь Ляо TL. Разработка надежных и настраиваемых пространственных структур с синтетическими генными цепями. Nucleic Acids Res. 2016; 1–15. - 45.
Пейн С., Ли Б., Цао И., Шеффер Д., Райзер М. Д., Ю Л. Временный контроль формирования самоорганизованного паттерна без градиентов морфогенов у бактерий. Mol Syst Biol. Издательская группа «Природа»; 2013; 9: 1–10. pmid: 24104480 - 46.
Марш JC, Bentley WE.Зондирование кворума и бактериальный перекрестный разговор в биотехнологии. Curr Opin Biotechnol. 2004; 15: 495–502. pmid: 15464383 - 47.
Ансальди М., Маролт Д., Стебе Т., Мандич-Мулек И., Дубнау Д. Специфическая активация систем восприятия кворума Bacillus вариантами изопренилированного феромона. Mol Microbiol. 2002; 44: 1561–1573. pmid: 12067344 - 48.
Чугани С.А., Уайтли М., Ли К.М., Д’Аргенио Д., Манойл С., Гринберг Е.П. QscR, модулятор синтеза кворум-чувствительного сигнала и вирулентности у синегнойной палочки.Proc Natl Acad Sci U S. A. 2001; 98: 2752–2757. pmid: 11226312 - 49.
Фридман Дж., Гор Дж. Экологическая системная биология: динамика взаимодействующих популяций. Curr Opin Syst Biol. Elsevier Ltd; 2017; 1: 114–121. - 50.
Hibbing ME, Fuqua C, Parsek MR, Peterson SB. Джунгли. Natl Rev Microbiol. 2010; 8: 15–25. - 51.
Кремер Н.Н., Филлипс Р., Ньюман Д.К., Бодиккер Дж.В. Прогнозирование влияния изменчивости промотора на регулирующие результаты. Научный представительИздательская группа «Природа»; 2015; 5: 18238. pmid: 26675057 - 52.
Guo X, Boedicker JQ. Вклад метаболических взаимодействий высокого порядка в глобальную активность четырехвидового микробного сообщества. PLoS Comput Biol. 2016; 12: 1–13. pmid: 27623159
Удобный для пользователя подход к оценке программ и эффективных общественных мероприятий для семей, подверженных риску бездомности, на JSTOR
Abstract
В этой статье показано, как удобная для пользователя оценка финансируемой из федерального бюджета программы профилактики бездомных с использованием исследовательского подхода и использования логической модели в качестве аналитической основы информировала множество заинтересованных сторон, включая членов Конгресса, других лиц, принимающих решения, и Семейного центра. практикующие.Целевой группой программы были семьи с очень низким доходом, которым грозило выселение из государственного жилья. Авторы обсуждают использованные методы, применение логической модели и результаты исследования по мере их развертывания в четыре этапа: (1) логическое моделирование как планирование программы, (2) концептуализация вмешательства, (3) определение процессов реализации и ( 4) определение диапазона результатов для клиента. Обсуждаются последствия для социальной политики, практики социальной работы и оценочных исследований.
Информация журнала
Социальная работа — ведущий журнал о профессии социальной работы. Это официальный журнал NASW, широко читаемый практиками, преподавателями и студентами, предоставляется всем членам в качестве членского преимущества. Социальная работа направлена на совершенствование практики и углубление знаний в области социальной работы и социального обеспечения. В его статьях можно по-новому взглянуть на сложившуюся практику, оценить новые методы и исследования, изучить текущие социальные проблемы и провести серьезный критический анализ проблем в профессии.Основной упор делается на социальную политику и решение серьезных человеческих проблем.
Информация об издателе
Oxford University Press — это отделение Оксфордского университета. Издание во всем мире способствует достижению цели университета в области исследований, стипендий и образования. OUP — крупнейшая в мире университетская пресса с самым широким глобальным присутствием. В настоящее время он издает более 6000 новых публикаций в год, имеет офисы примерно в пятидесяти странах и насчитывает более 5500 сотрудников по всему миру.Он стал известен миллионам людей благодаря разнообразной издательской программе, которая включает научные работы по всем академическим дисциплинам, библии, музыку, школьные и университетские учебники, книги по бизнесу, словари и справочники, а также академические журналы.
404 не найдено
404 не найдено
Запрошенный URL /~sbuss/researchweb/handbookii/chapterii.pdf не найден на этом сервере.
Наиболее частые причины этой ошибки:
- Вы неправильно ввели URL-адрес, к которому вы пытаетесь получить доступ.Тщательно проверьте орфографию, пунктуацию и чувствительность к регистру URL-адреса и повторите попытку.
- Файл или каталог, к которому вы пытаетесь получить доступ, больше не существует или был перемещен в другое место.
Если вам нужна помощь в разрешении этой проблемы, обратитесь к владельцу веб-страницы или веб-мастеру, как описано ниже.
Информацию о веб-сайтах класса можно найти в списке веб-сайтов класса на
http://www.math.ucsd.edu/resources/course-websites/.
Для других веб-страниц, пожалуйста, начните с веб-сайта верхнего уровня математического факультета UCSD по адресу
http: // www.math.ucsd.edu/.
Чтобы связаться с администраторами веб-сервера, отправьте электронное письмо по адресу
[email protected].
Чтобы мы могли должным образом устранить проблему, включите:
- Точный URL-адрес, который вы пытаетесь получить, указан в вашем веб-браузере:
REQUEST_URI = http://www.math.ucsd.edu/~sbuss/researchweb/handbookii/chapterii.pdf - Предыдущая ссылающаяся веб-страница или ссылка, которая привела вас к этому URL-адресу:
HTTP_REFERER = (нет) - Полное имя используемого вами веб-браузера, включая номер его версии:
HTTP_USER_AGENT = Mozilla / 5.