«Рисунок несуществующего животного»
1. Название:
методика «Рисунок несуществующего
животного» (РНЖ).
2. Автор:
Дукаревич М.З.
Контингент
испытуемых:
методика может использоваться для
обследования как детей, так и взрослых.Предмет
диагностики:
В терминах
рубрикатора:
индивидуально-целевые черты личности,
функциональные и эмоциональные
состояния.в терминах автора
методики:
черты личности, проблемы и социальные
установки испытуемого.
Операциональный
статус:
проективная техника изучения проективных
предпочтений.Сопряженные
практические задачи:
может использоваться как вспомогательный
диагностический прием в процессе
групповой психокоррекции, либо в
клиническом обследовании; рисунок
может быть использован как материал
для живого обсуждения в группе пациентов,
либо в диалоге «исследователь –
испытуемый»; в плане индивидуального
консультирования может использоваться
в качестве эвристической опоры для
направления беседы.
Компетентность
исполнителя:
проведение диагностики, обработка и
интерпретация данных может осуществляться
только специалистом.Комплектность,
состав методики:
существуют инструкции по проведению
и интерпретации результатов. По составу
данный тест – ориентировочный и как
единственный метод исследования обычно
не используется и требует объединения
с другими методами в качестве батарейного
инструмента исследования.Теоретические
положения: Метод
исследования построен на теории
психомоторной связи. Для регистрации
состояния психики используется
исследование моторики (в частности,
моторики рисующей доминантной руки –
зафиксированной в виде графического
следа движения, рисунка). Рисунок
правомерно отнести к области невербальной
(в данном случае – графической)
коммуникации. Важной характеристикой
невербального канала коммуникации
является его связь с эмоциями, играющими
определяющую роль в формировании
образов фантазии. Второй важной
характеристикой является связь рисунка
с метафорами речи. Одним из опорных
положений при интерпретации является
семантический параллелизм
образно-графического и вербально-метафорического
рядов. В самом общем виде процесс
интерпретации состоит в следующем: то,
что содержится в рисунке (местоположение,
детали и их взаимоотношения), буквально
переносится на личность рисовавшего
в качестве его метафорического описания.
Суждение о личности, как правило,
формируется в результате перевода с
языка метафорического описания на
привычный «профессиональный жаргон».
Пример: шипы – «личность с шипами» –
склонность к защитной агрессии; большие
уши – «человек с большими ушами» –
повышенная заинтересованность в
информации о себе.Оборудование:

 Второй важной
Второй важной 1. Белый или слегка
кремовый, не глянцевый лист бумаги.
Лист должен быть ориентирован
вертикально.
2. Простой карандаш
средней мягкости, неостро отточенный;
ручкой и фломастером рисовать нельзя.
11. Ход
работы:
Испытуемому предоставляется необходимое
оборудование. После этого испытуемому
предлагается инструкция следующего
содержания: «Придумайте и нарисуйте
несуществующее животное и назовите
его несуществующим названием». Далее
можно уточнить, что не желательно брать
животное из мультфильмов, т.к. оно уже
кем-то придумано; вымершие животные
тоже не подходят». После того, как рисунок
выполнен, и несуществующее животное
получило свое имя, предлагается выделить
основные параметры и провести
психологическую интерпретацию
изображенного на рисунке.
В заключение
работы на основе полученных материалов
составляется развернутая характеристика
психических состояний и свойств
личности.
Возможно групповое
обследование.
Аспекты анализа
рисунка:
Аспекты
анализа рисунка разделяются на формальные
и содержательные.
Формальный
анализ рисунка
К формальному
аспекту относятся:
а) семантика
расположения в пространстве,
б) графологические
признаки.
Дукаревич Майя Захаровна. Проективная методика «Несуществующие животные»
Похожие презентации:
Темперамент. Типы темперамента
Воображение и его виды
Вербальные и невербальные средства общения
Конфликты и способы их разрешения
Культурно-историческая концепция Л.С. Выготского
Кейсы (ситуации взаимодействия ребёнка и взрослого)
Гуманистическая психология
Анализ воздействия социальных сетей на формирование зависимого поведения у молодежи
Деловая коммуникация. Формы деловой коммуникации. (Лекция 2)
Технологии нейрокоррекции нарушений письменной речи младших школьников с тяжелыми нарушениями речи
1. Проективная методика: «Несуществующие животные»
2. Дукаревич Майя Захаровна
(14 мая 1925, Москва — 16 августа 2001, Москва)
3. Краткая биография
• Родилась в семье советского чиновника и дворянки. По дворянской
традиции до третьего класса девочка получала домашнее образование,
обучаясь гувернантками.
• С детства владела французским и немецким языками. Во время войны
училась на романо-германском факультете филологического
факультета. Была отчислена на пятом курсе.
• Продолжала самообразование, найдя своё призвание в психологии.
• В 1970-х гг. М. З. Дукаревич совместно с Ю. С. Савенко перевели и
адаптировали тест Роршаха и методику «Рисунок человека». Тогда же
была разработана и методика «Рисунок несуществующего животного».
Однако впервые под фамилией автора она была опубликована лишь в
1990-е гг
Инструкция:
Придумайте и нарисуйте
несуществующее животное и
назовите его несуществующим
названием.
многоголовка
животное, которое
служит домом для
людей
аура
АГРЕССИВНОСТЬ
НИЗКИЙ УРОВЕНЬ АГРЕССИВНОСТИ
Наиболее полную информацию об уровне агрессивности обследуемого дает сопоставление
классического варианта методики «Несуществующее животное» с дополнительным вариантом «Злое
животное».
ПОВЫШЕННАЯ АГРЕССИВНОСТЬ
Один из наиболее распространен-ных признаков повышенной агрессивности в рисунке
несуществующего животного — это наличие острых выступов и выростов, независимо от того, что они
изображают (рога, уши, щупальца, клешни). Примером такого рисунка может служить драморобот,
Примером такого рисунка может служить драморобот,
изображенный восьмилетним Артуром С.
Рисунки Володи С. (14 лет)
«Таким я стараюсь выглядеть (боксер), а такой я на самом деле (викон-горигозавр)».
СКЛОННОСТЬ К ВЕРБАЛЬНОЙ АГРЕССИИ
В рисунке несуществующего животного склонность к вербальной агрессии, как и в рисунке человека, выражается
в подчеркивании зубов. Так, Лена, 17 лет. по стандартной инструкции изобразила животное под названием
Весельчак.
БОЯЗНЬ АГРЕССИИ И ЗАЩИТНАЯ АГРЕССИЯ
Боязнь нападения приводит к стремлению защитить придуманное животное. В качестве защиты может быть
изображен панцирь, чешуя, броня, особо толстая кожа (ее может не быть на рисунке, но ее описывают в
рассказе). Очень широко распространено изображение игл, как у дикобраза, или колючек.
Ильи Р. 14 лет.
НЕВРОТИЧЕСКАЯ АГРЕССИЯ
Люда
«Питанием это чудовища являются земельные животные.
Своими длинными руками он достает их из-под земли. Также
представляет опасность для животных.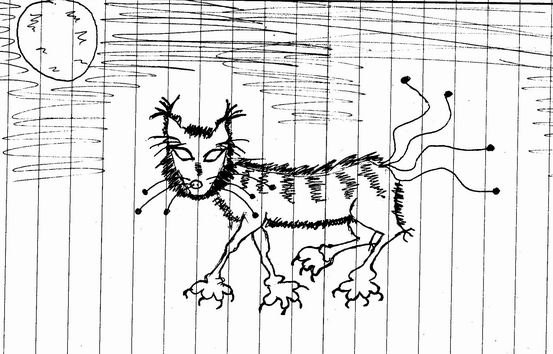 Оно высасывает из
Оно высасывает из
них кровь своими острыми жалами».
Грубое искажение формы глаз (в данном случае превращение их в агрессивные острия) — один из показателей
невротизации.
Если описание просто несуществующего животного сделано
вполне литературно, развернутыми фразами, то при
описании злого и страшного животного используются
рубленые, предельно упрощенные фразы; нарушается
согласование, появляются грубые ошибки внимания
(«питанием это чудовища являются…»).
РЕАКЦИЯ НА ЭМОЦИОНАЛЬНУЮ НАГРУЗКУ
Татьяна Н. по
стандартной
инструкции
изобразила животное
под названием «блин»
Аня К., 4 лет 8 месяцев
ПРИЗНАКИ ВОЗМОЖНОЙ ПСИХИЧЕСКОЙ ПАТОЛОГИИ.
Сергей 12 лет
Андрей Р. (15 лет.) «Получеловек»
«Мутант. Человек постядерной цивилизации с диградированным
мышлением. Видет почти обычный образ жизни человека, но с повадками
животного. Друзья — люди. Имеет неизмеримую мышечную массу». На
вопрос, в чем состоит его «несчастность», мальчик ответил: «Его вид его
огорчает. Не похож на людей, изменен. Люди к нему по-другому относятся».
Не похож на людей, изменен. Люди к нему по-другому относятся».
Затем был задан вопрос, в чем проявляются «повадки животного», о которых
он написал в рассказе. Андрей ответил: «Может на дерево залезть. Иногда
думает об одном, а потом переключается на другое. Забывает что-то».
English
Русский
Правила
Расширение суперсемейства основных фасилитаторов (MFS) для включения новых транспортеров, а также ферментов трансмембранного действия
[1] Saier MH Jr., Tran CV, Barabote RD, TCDB: База данных по классификации транспортеров для анализа мембранных транспортных белков и информации , Исследование нуклеиновых кислот, 34 (2006) D181–186. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[2] Saier MH Jr., Yen MR, Noto K, Tamang DG, Elkan C, База данных классификации переносчиков: последние достижения, Исследование нуклеиновых кислот, 37 (2009 г.)) Д274–278. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[3] Yee DC, Shlykov MA, Vastermark A, Reddy VS, Arora S, Sun EI, Saier MH Jr. , Рецептор, связанный с белком транспортер-опсин-G (TOG) надсемейство, FEBS J, 280 (2013) 5780–5800. [Бесплатная статья PMC] [PubMed] [Google Scholar]
, Рецептор, связанный с белком транспортер-опсин-G (TOG) надсемейство, FEBS J, 280 (2013) 5780–5800. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[4] Marger MD, Saier MH Jr., Основное суперсемейство трансмембранных посредников, которые катализируют унипорт, симпорт и антипорт, Trends Biochem Sci, 18 (1993) 13–20 . [PubMed] [Google Scholar]
[5] Pao SS, Paulsen IT, Saier MH Jr., Major facilitator superfamilitator, Microbiol Mol Biol Rev, 62 (19)98) 1–34. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[6] Reddy VS, Saier MH Jr., BioV Suite — набор программ для изучения эволюции транспортных белков, Febs j, 279 (2012) 2036– 2046. [Статья бесплатно PMC] [PubMed] [Google Scholar]
[7] Park JH, Saier MH Jr., Филогенетическая характеристика семейства белков трансмембранных каналов MIP, J Membr Biol, 153 (1996) 171–180. [PubMed] [Google Scholar]
[8] Морган Дж. Л., Струмилло Дж., Циммер Дж., Кристаллографический снимок синтеза целлюлозы и мембранной транслокации, Nature, 49.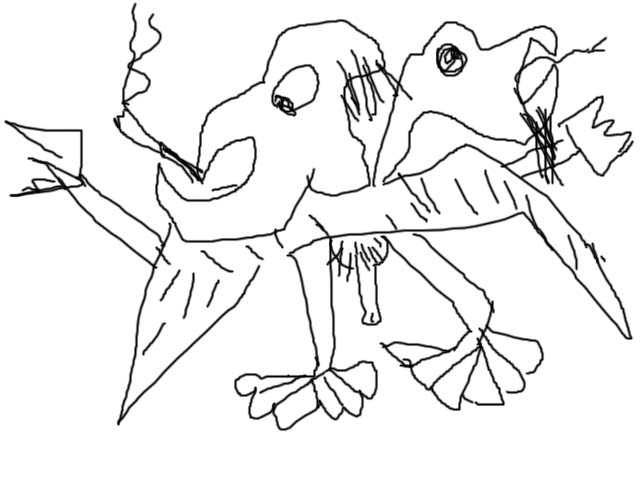 3 (2013) 181–186. [Статья бесплатно PMC] [PubMed] [Google Scholar]
3 (2013) 181–186. [Статья бесплатно PMC] [PubMed] [Google Scholar]
[9] Saier MH Jr., Компьютерный анализ последовательностей транспортных белков: сбор данных о функции, структуре, биогенезе и эволюции, Microbiol Rev, 58 (1994) 71–93. [Статья бесплатно PMC] [PubMed] [Google Scholar]
[10] Saier MH Jr., Функционально-филогенетическая система классификации трансмембранных переносчиков растворенных веществ, Microbiol Mol Biol Rev, 64 (2000) 354–411. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[11] Saier MH Jr., Reddy VS, Tsu BV, Ahmed MS, Li C, Moreno-Hagelsieb G, База данных классификации переносчиков (TCDB): последние достижения, Исследование нуклеиновых кислот, 44 (2016) D372–379. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[12] Редди В.С., Шлыков М.А., Кастильо Р., Сун Э.И., Сайер М.Х. мл., Новый взгляд на главное суперсемейство фасилитаторов (MFS), FEBS J, 279 (2012) 2022–2035 гг. [PMC free article] [PubMed] [Google Scholar]
[13] Wang B, Dukarevich M, Sun EI, Yen MR, Saier MH Jr. , Мембранные портеры АТФ-связывающих кассетных транспортных систем полифилетичны, J Membr Biol, 231 (2009 г.)) 1–10. [PubMed] [Google Scholar]
, Мембранные портеры АТФ-связывающих кассетных транспортных систем полифилетичны, J Membr Biol, 231 (2009 г.)) 1–10. [PubMed] [Google Scholar]
[14] Hirai T, Heymann JA, Maloney PC, Subramaniam S, Структурная модель 12-спиральных транспортеров, принадлежащих к главному надсемейству фасилитаторов, Journal of bacteriology, 185 (2003) 1712–1718. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[15] Vastermark A, Saier MH, Major Facilitator Superfamily (MFS) развилась без реаранжировки 3-трансмембранных сегментов, Proceedings of the National Academy of Sciences of the United States of Америка, 111 (2014) E1162–1163. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[16] Ян Н., Структурные достижения для транспортеров главного суперсемейства фасилитаторов (MFS), Trends Biochem Sci, 38 (2013) 151–159. [PubMed] [Google Scholar]
[17] Мораес Т.Ф., Райтмайер Р.А., Мембранные транспортные метаболоны, Biochimica et biophysica acta, 1818 (2012) 2687–2706. [PubMed] [Google Scholar]
[18] Ян Н. , Структурная биология транспортеров надсемейства главных помощников, Ежегодный обзор биофизики, 44 (2015) 257–283. [PubMed] [Google Scholar]
, Структурная биология транспортеров надсемейства главных помощников, Ежегодный обзор биофизики, 44 (2015) 257–283. [PubMed] [Google Scholar]
[19] Kaback HR, Хемиосмотический механизм симпорта, Труды Национальной академии наук Соединенных Штатов Америки, 112 (2015) 1259–1264. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[20] Zhang XC, Zhao Y, Heng J, Jiang D, Механизмы энергетического сцепления транспортеров MFS, Наука о белках: публикация Белкового общества, 24 (2015) 1560–1579 гг. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[21] Wisedchaisri G, Park MS, Iadanza MG, Zheng H, Gonen T, Связанный с протоном транспорт сахара в прототипе главного облегчающего белка суперсемейства XylE, Nat Commun, 5 (2014) 4521. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[22] Сайер М.Х. младший, Битти Дж.Т., Гоффо А., Харли К.Т., Хейне В.Х., Хуан С.К., Джек Д.Л., Ян П.С., Лью К., Лю Дж., Пао С.С., Полсен ИТ, Ценг Т.Т., Вирк П.С., главное суперсемейство помощников, Журнал молекулярной микробиологии и биотехнологии, 1 (1999) 257–279. [PubMed] [Google Scholar]
[PubMed] [Google Scholar]
[23] Winkler HH, Neuhaus HE, Немитохондриальный транспорт АТФ, Trends Biochem Sci, 24 (1999) 64–68. [PubMed] [Google Scholar]
[24] Trentmann O, Jung B, Neuhaus HE, Haferkamp I, Немитохондриальные транспортеры АТФ/АДФ принимают фосфат в качестве третьего субстрата, Журнал биологической химии, 283 (2008) 36486–36493. [Статья бесплатно PMC] [PubMed] [Google Scholar]
[25] Ньюстед С., Молекулярные исследования транспорта пептидов, связанных с протонами, в семействе переносчиков олигопептидов PTR, Biochimica et biophysica acta, 1850 (2015) 488–499. [Статья бесплатно PMC] [PubMed] [Google Scholar]
[26] Ньюстед С., На пути к структурному пониманию транспорта лекарств и пептидов в семействе протонзависимых олигопептидных транспортеров (POT), Biochem Soc Trans, 39 (2011) 1353 –1358. [PubMed] [Академия Google]
[27] Мазерли Л.Х., Уилсон М.Р., Хоу З., Основные вспомогательные переносчики фолиевой кислоты, переносчик растворенных веществ 19A1 и переносчик растворенных веществ 46A1: биология и роль в антифолатной химиотерапии рака, Метаболизм и расположение лекарств: биологическая судьба химических веществ, 42 (2014 г. ) ) 632–649. [Статья бесплатно PMC] [PubMed] [Google Scholar]
) ) 632–649. [Статья бесплатно PMC] [PubMed] [Google Scholar]
[28] Flintoff WF, Williams FM, Sadlish H, Область между трансмембранными доменами 1 и 2 восстановленного носителя фолиевой кислоты образует часть кармана, связывающего субстрат, The Journal биологической химии, 278 (2003) 40867–40876. [PubMed] [Академия Google]
[29] Хонг М., Критические домены в последовательности полипептидов, переносящих органические анионы человека, Текущий метаболизм лекарств, 15 (2014) 265–270. [PubMed] [Google Scholar]
[30] Hagenbuch B, Stieger B, Надсемейство транспортеров SLCO (ранее SLC21), Молекулярные аспекты медицины, 34 (2013) 396–412. [PMC free article] [PubMed] [Google Scholar]
[31] Цигельный И.Ф., Ковальский Д., Кузнецова В.Л., Балинский О., Шариков Ю., Бхатнагар В., Нигам С.К., Конформационные изменения мультиспецифического транспортера транспортера органических анионов 1 (OAT1 /SLC22A6) предполагает молекулярный механизм начальных стадий транспорта лекарств и метаболитов, Cell biochemistry and biophysics, 61 (2011) 251–259. [PubMed] [Google Scholar]
[PubMed] [Google Scholar]
[32] Zhu C, Nigam KB, Date RC, Bush KT, Springer SA, Saier MH Jr., Wu W, Nigam SK, Эволюционный анализ и классификация OAT, OCT, OCTN и Другие транспортеры SLC22: значение структуры и функции и анализ мотивов последовательности, PloS one, 10 (2015) e0140569. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[33] Tazoe Y, Hayashi H, Tsuboi S, Shioura T, Matsuyama T, Yamada H, Hirai K, Tsuji D, Inoue K, Sugiyama T, Itoh K, Снижение уровня экспрессии гена переносчика 1 фолиевой кислоты коррелирует с эффективностью метотрексата у японских пациентов с ревматоидным артритом, Метаболизм лекарственных средств и фармакокинетика, 30 (2015) 227–230. [PubMed] [Академия Google]
[34] Eudes A, Kunji ER, Noiriel A, Klaus SM, Vickers TJ, Beverley SM, Gregory JF 3rd, Hanson AD, Идентификация критических для транспорта остатков в переносчике фолиевой кислоты из семейства переносчиков фолата-биоптерина (FBT). , Журнал биологической химии, 285 (2010) 2867–2875. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
[35] Vastermark A, Driker A, Li J, Saier MH Jr., Консервативное движение TMS11 между закрытыми конформациями LacY и XylE главного надсемейства фасилитаторов предполагает подобное шарнирный механизм, Белки, 83 (2015) 735–745. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[36] Wong WC, Maurer-Stroh S, Eisenhaber F, Не все трансмембранные спирали рождаются одинаковыми: К расширению концепции гомологии последовательностей на мембранные белки, Biol Direct, 6 (2011) 57. [Статья бесплатно PMC] [PubMed] [Google Scholar]
[37] Wong WC, Maurer-Stroh S, Schneider G, Eisenhaber F, Трансмембранная спираль: простая или сложная, Исследование нуклеиновых кислот, 40 (2012) W370–375. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[38] El-Gebali S, Mistry J, Bateman A, Eddy SR, Luciani A, Potter SC, Qureshi M, Richardson LJ, Salazar GA, Smart A, Sonnhammer ELL, Хирш Л., Паладин Л., Пиовесан Д., Tosatto SCE, Финн Р. Д., База данных семейств белков Pfam в 2019 г., Исследование нуклеиновых кислот, 47 (2019) D427–D432. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Д., База данных семейств белков Pfam в 2019 г., Исследование нуклеиновых кислот, 47 (2019) D427–D432. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[39] Zhai Y, Saier MH Jr., Интернет-программа для прогнозирования средней гидропатии, средней амфипатичности и среднего сходства множественно выровненных гомологичных белков, Journal of молекулярная микробиология и биотехнология, 3 (2001) 285–286. [PubMed] [Google Scholar]
[40] Медрано-Сото А., Морено-Хагельсиб Г., Маклафлин Д., Йе З.С., Хендарго К.Дж., Сайер М.Х. мл., Биоинформатическая характеристика аноктаминового надсемейства Са2+-активируемых ионных каналов и липидов Scramblases, PloS one, 13 (2018) e0192851. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[41] Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ, Gapped BLAST и PSI-BLAST: новое поколение программ поиска в базе данных белков, Исследование нуклеиновых кислот, 25 (1997) 3389–3402. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[42] Fu L, Niu B, Zhu Z, Wu S, Li W, CD-HIT: ускорено для кластеризации данных секвенирования следующего поколения, Bioinformatics (Oxford, Англия), 28 (2012) 3150–3152. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
[43] Пирсон В. Р., Поиск библиотек белковых последовательностей: сравнение чувствительности и селективности алгоритмов Смита-Уотермана и FASTA, Genomics, 11 (1991) 635–650. [PubMed] [Google Scholar]
[44] Wong WC, Maurer-Stroh S, Eisenhaber F, Более 1001 проблемы с базами данных белковых доменов: трансмембранные области, сигнальные пептиды и проблема гомологии последовательностей, PLoS Comput Biol, 6 ( 2010) e1000867. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[45] Katoh K, Standley DM, Программное обеспечение для множественного выравнивания последовательностей MAFFT, версия 7: улучшения производительности и удобства использования, Mol Biol Evol, 30 (2013) 772–780. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[46] Capella-Gutierrez S, Silla-Martinez JM, Gabaldon T, trimAl: инструмент для автоматической обрезки выравнивания в крупномасштабном филогенетическом анализе, Bioinformatics (Oxford, England), 25 (2009) 1972–1973. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
[47] Zhai Y, Saier MH Jr., Веб-программа (WHAT) для одновременного прогнозирования гидропатии, амфипатии, вторичной структуры и трансмембранной топологии для одного белковая последовательность, Журнал молекулярной микробиологии и биотехнологии, 3 (2001) 501–502. [PubMed] [Академия Google]
[48] Tusnady GE, Simon I, Сервер предсказания трансмембранной топологии HMMTOP, Bioinformatics (Oxford, England), 17 (2001) 849–850. [PubMed] [Google Scholar]
[49] Бернсель А., Виклунд Х., Хеннердал А., Элофссон А., TOPCONS: консенсусное предсказание топологии мембранных белков, Исследование нуклеиновых кислот, 37 (2009) W465–468. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[50] Редди А., Чо Дж., Линг С., Редди В., Шлыков М., Сайер М.Х., Надежность девяти программ топологических прогнозов и их применение к интегральному мембранному каналу и белки-носители, Журнал молекулярной микробиологии и биотехнологии, 24 (2014) 161–190. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
[51] Ломизе М.А., Погожева И.Д., Джу Х., Мосберг HI, Ломизе А.Л., база данных OPM и веб-сервер PPM: ресурсы для позиционирования белков в мембранах, Nucleic исследования кислот, 40 (2012) D370–376. [Статья бесплатно PMC] [PubMed] [Google Scholar]
[52] Tusnady GE, Dosztanyi Z, Simon I, PDB_TM: отбор и мембранная локализация трансмембранных белков в банке данных белков, Исследование нуклеиновых кислот, 33 (2005) D275 –278. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[53] Фришман Д., Аргос П., Присвоение вторичной структуры белка на основе знаний, Proteins, 23 (1995) 566–579. [PubMed] [Google Scholar]
[54] Winn MD, Ballard CC, Cowtan KD, Dodson EJ, Emsley P, Evans PR, Keegan RM, Krissinel EB, Leslie AG, McCoy A, McNicholas SJ, Murshudov GN, Pannu NS , Potterton EA, Powell HR, Read RJ, Vagin A, Wilson KS, Обзор пакета CCP4 и текущих разработок, Acta Crystallogr D Biol Crystallogr, 67 (2011) 235–242. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
[55] Krissinel E, Henrick K, Согласование вторичной структуры (SSM), новый инструмент для быстрого выравнивания структуры белка в трех измерениях, Acta Crystallogr D Biol Crystallogr, 60 (2004) 2256–2268. [PubMed] [Google Scholar]
[56] Zhang Y, Skolnick J, TM-align: алгоритм выравнивания структуры белка на основе TM-score, Nucleic acid research, 33 (2005) 2302–2309. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[57] Yen MR, Choi J, Saier MH Jr, Биоинформатический анализ трансмембранного транспорта: новое программное обеспечение для определения филогении, топологии и эволюции белков, Журнал молекулярной микробиологии и биотехнологии, 17 (2009)) 163–176. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[58] Eddy SR, Accelerated Profile HMM Searches, PLoS Comput Biol, 7 (2011) e1002195. [Статья бесплатно PMC] [PubMed] [Google Scholar]
[59] Valdes R, Elferich J, Shinde U, Landfear SM, Идентификация внутриклеточных ворот для члена семейства уравновешивающих нуклеозидных транспортеров (ENT), The Journal of биологическая химия, 289 (2014) 8799–8809. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
[60] Baldwin SA, Beal PR, Yao SY, King AE, Cass CE, Young JD, Семейство уравновешивающих нуклеозидных переносчиков, SLC29, Pflugers Archiv: Европейский журнал физиологии, 447 (2004) 735–743. [PubMed] [Google Scholar]
[61] Harley KT, Djordjevic GM, Tseng TT, Saier MH, Гомологи слитых с мембраной белков в грамположительных бактериях, Mol Microbiol, 36 (2000) 516–517. [PubMed] [Google Scholar]
[62] Harley KT, Saier MH Jr., Новое вездесущее семейство предполагаемых транспортеров оттока, Журнал молекулярной микробиологии и биотехнологии, 2 (2000) 195–198. [PubMed] [Google Scholar]
[63] Ван Дайк Т.К., Темплтон Л.Дж., Кантера К.А., Шарп П.Л., Сариаслани Ф.С., Характеристика эффлюксного насоса Escherichia coli AaeAB: метаболический предохранительный клапан?, Журнал бактериологии, 186 (2004 г.) ) 7196–7204. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[64] Yeh KY, Yeh M, Glass J, Взаимодействие между ферропортином и гефестином в энтероцитах крыс снижается после приема железа, Gastroenterology, 141 (2011) 292–299, 299. e291. [PubMed] [Google Scholar]
e291. [PubMed] [Google Scholar]
[65] Madejczyk MS, Ballatori N, Переносчик железа ферропортин также может функционировать как экспортер марганца, Biochimica et biophysica acta, 1818 (2012) 651–657. [Статья бесплатно PMC] [PubMed] [Google Scholar]
[66] Delaby C, Pilard N, Puy H, Canonne-Hergaux F, Последовательная регуляция экспрессии ферропортина после эритрофагоцитоза в мышиных макрофагах: ранняя индукция мРНК гемом с последующей экспрессия железозависимого белка, The Biochemical Journal, 411 (2008) 123–131. [PubMed] [Академия Google]
[67] Mitchell CJ, Shawki A, Ganz T, Nemeth E, Mackenzie B, Функциональные свойства человеческого ферропортина, экспортера клеточного железа, реагирующего также с кобальтом и цинком, Американский журнал физиологии. Клеточная физиология, 306 (2014) C450–459. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[68] Танигути Р., Като Х.Э., Фонт Дж., Дешпанде К.Н., Вада М., Ито К., Ишитани Р., Джормакка М., Нуреки О., обращенные наружу и внутрь структуры предполагаемого бактериального переносчика переходных металлов с гомологией ферропортину, Nat Commun, 6 (2015) 8545. [Статья бесплатно PMC] [PubMed] [Google Scholar]
[Статья бесплатно PMC] [PubMed] [Google Scholar]
[69] Nomura N, Verdon G, Kang HJ, Shimamura T, Nomura Y, Sonoda Y, Hussien SA, Qureshi AA, Coincon M, Sato Y, Abe H, Nakada-Nakura Y, Hino T, Arakawa T, Kusano -Арай О., Иванари Х., Мурата Т., Кобаяши Т., Хамакубо Т., Касахара М., Ивата С., Дрю Д., Структура и механизм переносчика фруктозы млекопитающих GLUT5, Nature, 526 (2015) 397–401. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[70] Йонезава А., Масуда С., Кацура Т., Инуи К., Идентификация и функциональная характеристика нового переносчика рибофлавина человека и крысы, RFT1, Американский журнал физиологии. Клеточная физиология, 295 (2008) C632–641. [PubMed] [Google Scholar]
[71] Powers HJ, Рибофлавин (витамин B-2) и здоровье, Американский журнал клинического питания, 77 (2003) 1352–1360. [PubMed] [Google Scholar]
[72] Yamamoto S, Inoue K, Ohta KY, Fukatsu R, Maeda JY, Yoshida Y, Yuasa H, Идентификация и функциональная характеристика переносчика рибофлавина крысы 2, Журнал биохимии, 145 (2009 г. ) ) 437–443. [PubMed] [Google Scholar]
) ) 437–443. [PubMed] [Google Scholar]
[73] Ernst CM, Peschel A, Устойчивость к противомикробным пептидам широкого спектра действия посредством MprF-опосредованного аминоацилирования и переворачивания фосфолипидов, Mol Microbiol, 80 (2011) 290–299. [PubMed] [Google Scholar]
[74] Ernst CM, Staubitz P, Mishra NN, Yang SJ, Hornig G, Kalbacher H, Bayer AS, Kraus D, Peschel A, Бактериальный белок устойчивости к дефенсину MprF состоит из разделяемых доменов для лизинилирование липидов и отталкивание противомикробных пептидов, PLoS Pathog, 5 (2009) e1000660. [Статья бесплатно PMC] [PubMed] [Google Scholar]
[75] Ernst CM, Kuhn S, Slavetinsky CJ, Krismer B, Heilbronner S, Gekeler C, Kraus D, Wagner S, Peschel A, The lipid-modifier Multiple peptide фактор резистентности представляет собой олигомер, состоящий из отдельных взаимодействующих субъединиц синтазы и флиппазы, MBio, 6 (2015). [Бесплатная статья PMC] [PubMed] [Google Scholar]
[76] Моцик А., Нечуштан Х., Фу С. Ю., Разин Э., Неканоническая роль лизил-тРНК-синтетазы в здоровье и болезнях, Тенденции в молекулярной медицине, 19 (2013) 726–731. [PubMed] [Google Scholar]
Ю., Разин Э., Неканоническая роль лизил-тРНК-синтетазы в здоровье и болезнях, Тенденции в молекулярной медицине, 19 (2013) 726–731. [PubMed] [Google Scholar]
[77] Каменский Ю., Лю В., Цай А.Л., Кулмац Р.Дж., Палмер Г. Осевое лигирование и стехиометрия гемовых центров в цитохроме b561 надпочечников, Биохимия, 46 (2007) 8647–8658. [Статья бесплатно PMC] [PubMed] [Google Scholar]
[78] Nakanishi N, Takeuchi F, Tsubaki M, Механизм гистидинового цикла для согласованного переноса протонов/электронов от аскорбата к цитозольному центру гема b цитохрома b561: уникальный аппарат для биологического трансмембранного переноса электронов, Журнал биохимии, 142 (2007) 553–560. [PubMed] [Академия Google]
[79] Liu W, Rogge CE, da Silva GF, Shinkarev VP, Tsai AL, Kamensky Y, Palmer G, Kulmacz RJ, His92 и His110 избирательно воздействуют на разные гемовые центры цитохрома b(561) надпочечников, Biochimica et biophysica acta , 1777 (2008) 1218–1228. [Статья бесплатно PMC] [PubMed] [Google Scholar]
[80] Lu P, Ma D, Yan C, Gong X, Du M, Shi Y, Структура и механизм эукариотической трансмембранной аскорбат-зависимой оксидоредуктазы, Труды Национальная академия наук Соединенных Штатов Америки, 111 (2014) 1813–1818.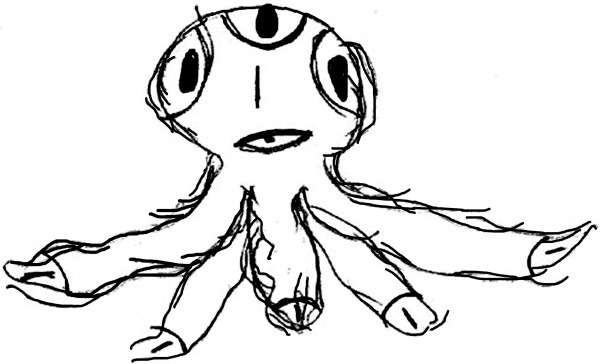 [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
[81] Lu Q, Lu JP, Li XD, Liu XH, Min H, Lin FC, Magnaporthe oryzae Ген MTP1 кодирует трансмембранный белок типа III, участвующий в конидиации и прорастании конидий, J Zhejiang Univ Sci B, 9 (2008) 511–519. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[82] Pao GM, Wu LF, Johnson KD, Hofte H, Chrispeels MJ, Sweet G, Sandal NN, Saier MH Jr., Эволюция интегрального семейства MIP мембранные транспортные белки, Mol Microbiol, 5 (1991) 33–37. [PubMed] [Академия Google]
[83] Абаскал Ф., Ирисарри И., Зардоя Р., Разнообразие и эволюция внутренних мембранных белков, Biochimica et biophysica acta, 1840 (2014) 1468–1481. [PubMed] [Google Scholar]
[84] Palmieri F, Семейство митохондриальных транспортеров SLC25: идентификация, свойства и физиопатология, Молекулярные аспекты медицины, 34 (2013) 465–484. [PubMed] [Google Scholar]
[85] Pebay-Peyroula E, Dahout-Gonzalez C, Kahn R, Trezeguet V, Lauquin GJ, Brandolin G, Структура митохондриального переносчика АДФ/АТФ в комплексе с карбоксиатрактилозидом, Nature, 426 ( 2003) 39–44. [PubMed] [Google Scholar]
[PubMed] [Google Scholar]
[86] Staubitz P, Neumann H, Schneider T, Wiedemann I, Peschel A, MprF-опосредованный биосинтез лизилфосфатидилглицерола, важной детерминанты устойчивости стафилококков к дефензину, FEMS Microbiol Lett, 231 (2004) 67–71. [PubMed] [Google Scholar]
[87] Andra J, Goldmann T, Ernst CM, Peschel A, Gutsmann T, Фактор множественной устойчивости к пептидам (MprF), опосредованный Устойчивость Staphylococcus aureus к противомикробным пептидам совпадает с модулированным взаимодействием пептидов с искусственные мембраны, содержащие лизилфосфатидилглицерин, Журнал биологической химии, 286 (2011) 18692–18700. [PMC free article] [PubMed] [Google Scholar]
[88] Slavetinsky CJ, Peschel A, Ernst CM, аланил-фосфатидилглицерин и лизил-фосфатидилглицерин транслоцируются одними и теми же флиппазами MprF и обладают сходной способностью защищать от антибиотика даптомицина. in Staphylococcus aureus, Antimicrob Agents Chemother, 56 (2012) 3492–3497. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[89] Hebecker S, Krausze J, Hasenkampf T, Schneider J, Groenewold M, Reichelt J, Jahn D, Heinz DW, Moser J, Structures of two bacterial Resistance Factors опосредуя тРНК-зависимое аминоацилирование фосфатидилглицерина лизином или аланином, Proceedings of the National Academy of Sciences of the United States of America, 112 (2015) 10691–10696. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
[90] Asard H, Barbaro R, Trost P, Berczi A, Cytochromes b561: аскорбат-опосредованный трансмембранный перенос электронов, Antioxid Redox Signal, 19 (2013) 1026 –1035. [Статья бесплатно PMC] [PubMed] [Google Scholar]
[91] Berczi A, Zimanyi L, Трансмембранные белки цитохрома b561: структурная информация и биологическая функция, Curr Protein Pept Sci, 15 (2014) 745–760. [PubMed] [Google Scholar]
[92] Lane DJ, Bae DH, Merlot AM, Sahni S, Richardson DR, Дуоденальный цитохром b (DCYTB) в метаболизме железа: обновленная информация о функции и регуляции, Nutrients, 7 (2015) 2274–2296. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Надсемейство аминокислот-полиаминов-органо-органов — Полный текст — Журнал молекулярной микробиологии и биотехнологии 2012, Vol. 22, No. 2
Показано, что суперсемейство аминокислот-полиаминов-органокатионитов (АРС) включает пять признанных семейств, четыре из которых специфичны для аминокислот и их производных. Недавние рентгеноструктурные данные высокого разрешения показали, что четыре дополнительных семейства транспортеров (BCCT, TC № 2.A.15; SSS, 2.A.21; NSS, 2.A.22; и NCS1, 2.A. .39), транспортирующие широкий спектр растворенных веществ, имеют достаточно схожие складки, чтобы предположить общее эволюционное происхождение. Мы использовали установленные статистические методы, основанные на сходстве последовательностей, чтобы показать, что эти семейства на самом деле являются членами суперсемейства APC. Мы также идентифицируем два дополнительных семейства (NCS2, 2.A.40; SulP, 2.A.53) как члены этого суперсемейства. Повторяющиеся последовательности, каждая из которых имеет пять трансмембранных α-спиральных сегментов и возникает в результате древних внутригенных дупликаций, продемонстрированы для всех этих семейств, что еще больше подтверждает вывод о гомологии. Надсемейство APC, по-видимому, является вторым по величине надсемейством вторичных носителей, самым большим из которых является надсемейство главных фасилитаторов (MFS).
Недавние рентгеноструктурные данные высокого разрешения показали, что четыре дополнительных семейства транспортеров (BCCT, TC № 2.A.15; SSS, 2.A.21; NSS, 2.A.22; и NCS1, 2.A. .39), транспортирующие широкий спектр растворенных веществ, имеют достаточно схожие складки, чтобы предположить общее эволюционное происхождение. Мы использовали установленные статистические методы, основанные на сходстве последовательностей, чтобы показать, что эти семейства на самом деле являются членами суперсемейства APC. Мы также идентифицируем два дополнительных семейства (NCS2, 2.A.40; SulP, 2.A.53) как члены этого суперсемейства. Повторяющиеся последовательности, каждая из которых имеет пять трансмембранных α-спиральных сегментов и возникает в результате древних внутригенных дупликаций, продемонстрированы для всех этих семейств, что еще больше подтверждает вывод о гомологии. Надсемейство APC, по-видимому, является вторым по величине надсемейством вторичных носителей, самым большим из которых является надсемейство главных фасилитаторов (MFS). Хотя топология членов суперсемейства APC отличается от топологии MFS, оба семейства, по-видимому, произошли от общей предковой шпилечной структуры 2 TMS, которая подверглась внутригенному трипликации с последующей потерей TMS в семействе APC, чтобы дать повтор единицы, характерные для этих двух надсемейств.
Хотя топология членов суперсемейства APC отличается от топологии MFS, оба семейства, по-видимому, произошли от общей предковой шпилечной структуры 2 TMS, которая подверглась внутригенному трипликации с последующей потерей TMS в семействе APC, чтобы дать повтор единицы, характерные для этих двух надсемейств.
Введение
На протяжении многих лет наша исследовательская группа идентифицировала и расширила суперсемейства трансмембранных белков-транспортеров, зарегистрированных в Базе данных классификации транспортеров [TCDB; www.tcdb.org; Сайер и др., 2006 г.; Сайер и др., 2009]. Эта база данных относит белки к суперсемействам и их соответствующим семействам или подсемействам, одновременно интегрируя новое биоинформатическое программное обеспечение и соответствующие ресурсы [Reddy and Saier, 2012]. Важность идентификации и классификации белковых семейств и суперсемейств подчеркивается тем фактом, что структурные, функциональные и механистические данные для транспортеров могут быть экстраполированы с одного белка на другой, если и только если было показано, что они связаны общим потомком. Лам и др., 2011; Сайер, 19 лет96, 1999, 2000].
Лам и др., 2011; Сайер, 19 лет96, 1999, 2000].
Основное внимание нашей лаборатории биоинформатики уделялось обновлению и разработке новых компьютерных программ, помогающих в идентификации и расширении белковых суперсемейств [Chang et al., 2004; Редди и Сайер, 2012]. Эти программы сыграли важную роль в установлении гомологии среди белков, имеющих общее происхождение. С каждым годом мы становимся все более опытными в надежном обнаружении и количественной оценке все более отдаленных филогенетических отношений. Параллельно мы совершенствуем искусство выявления и оценки эволюционных путей, которые привели к возникновению этих белков.
Надсемейство аминокислот-полиаминов-органокатион (APC) представляет собой большое надсемейство, которое на основе недавних трехмерных структурных анализов [Chan et al., 2010; Фанг и др., 2009 г.; Гао и др., 2010 г.; Джекельманн и др., 2011 г.; Kowalczyk et al., 2011] оказывается намного больше, чем считалось ранее. Ранее нами был установлен статус надсемейства надсемейства APC [Chang et al. , 2004; Джек и др., 2000; Young et al., 1999], но пять семейств, которые тогда были признаны для этого надсемейства, включали только одно из четырех семейств, которые, как предполагалось, были родственными на основании недавних трехмерных структурных данных.
, 2004; Джек и др., 2000; Young et al., 1999], но пять семейств, которые тогда были признаны для этого надсемейства, включали только одно из четырех семейств, которые, как предполагалось, были родственными на основании недавних трехмерных структурных данных.
Пять исходных семейств: (1) семейство APC (TC № 2.A.3), (2) семейство пермеаз аминокислот/ауксинов (AAAP; TC № 2.A.18), (3 ) семейство аланиновых или глицин:катионных симпортеров (AGCS; TC No. 2.A.25), (4) семейство катион-хлоридных котранспортеров (CCC; TC No. 2.A.30) и (5) гидроксильных / семейство пермеаз ароматических аминокислот (НАААР; ТК № 2.А.42) (таблица 1). Теперь мы представляем статистические результаты, основанные только на данных первичной последовательности, демонстрирующие гомологию ранее существовавших членов надсемейства APC со следующими шестью новыми семействами: (1) транспортер бетаина/карнитина/холина (BCCT; TC No. 2.A.15). ) семейство, (2) семейство растворенное вещество: симпортер натрия (SSS; ТК № 2. A.21), (3) семейство нейротрансмиттер: симпортер натрия (NSS; ТК № 2.A.22), (4) азотистое основание: катионный симпортер-1 (NCS1; ТК № 2.А.39) семейство, (5) семейство нуклеооснование: катионный симпортер-2 (NCS2; ТК № 2.А.40) и (6) семейство сульфатных пермеаз (SulP; ТК № 2.А.53) (табл. 1). ). Все эти семейства включают членов, которые демонстрируют 5 дуплицированных повторяющихся последовательностей TMS, хотя на их N- и C-концах могут существовать дополнительные трансмембранные α-спиральные сегменты (TMS). Мы приводим доказательства того, что единица из 5 повторов TMS возникла в результате внутригенного размножения, где первичная единица повтора включала только 2 TMS. Таким образом, путь появления всех членов надсемейства APC выглядит следующим образом:
A.21), (3) семейство нейротрансмиттер: симпортер натрия (NSS; ТК № 2.A.22), (4) азотистое основание: катионный симпортер-1 (NCS1; ТК № 2.А.39) семейство, (5) семейство нуклеооснование: катионный симпортер-2 (NCS2; ТК № 2.А.40) и (6) семейство сульфатных пермеаз (SulP; ТК № 2.А.53) (табл. 1). ). Все эти семейства включают членов, которые демонстрируют 5 дуплицированных повторяющихся последовательностей TMS, хотя на их N- и C-концах могут существовать дополнительные трансмембранные α-спиральные сегменты (TMS). Мы приводим доказательства того, что единица из 5 повторов TMS возникла в результате внутригенного размножения, где первичная единица повтора включала только 2 TMS. Таким образом, путь появления всех членов надсемейства APC выглядит следующим образом:
Таблица 1
Семейства в суперсемействе APC
Методы
Анализ выравнивания последовательностей
Последовательности FASTA, представляющие членов семейства APC (TC № 2.A.3), и каждый из вновь добавленные семьи в пределах Суперсемейство APC было собрано из TCDB [www.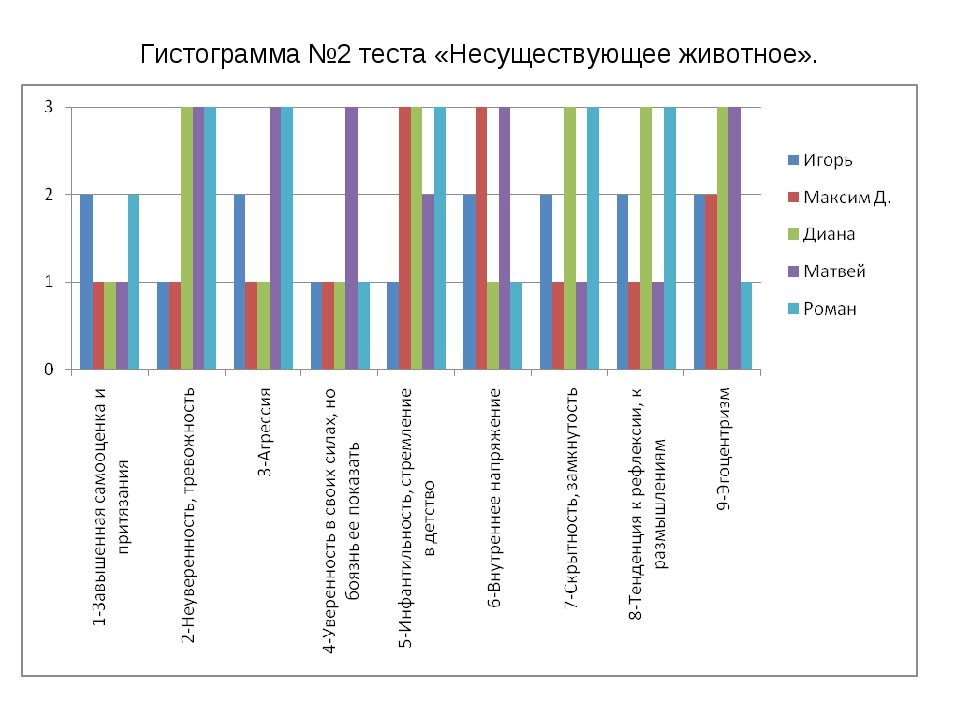 tcdb.org; Сайер и др., 2009]. Базовый инструмент локального поиска выравнивания (BLAST) [Altschul et al., 1990; Altschul et al., 1997] с использованием PSI-BLAST с двумя итерациями был выполнен на белках, представляющих каждое семейство, для создания семи списков гомологичных белков. В совокупности для анализа было собрано более 5600 белков из базы данных белков NR Национального центра биотехнологической информации (NCBI). Более 5000 белков были сохранены для дальнейшего исследования после удаления избыточности и белков, которые были по крайней мере 95% идентичны сохраненному белку с использованием модифицированной программы CD-Hit [Li and Godzik, 2006; Ли и др., 2001, 2002; Сайер и др., 2009 г.; Йен и др., 2009].
tcdb.org; Сайер и др., 2009]. Базовый инструмент локального поиска выравнивания (BLAST) [Altschul et al., 1990; Altschul et al., 1997] с использованием PSI-BLAST с двумя итерациями был выполнен на белках, представляющих каждое семейство, для создания семи списков гомологичных белков. В совокупности для анализа было собрано более 5600 белков из базы данных белков NR Национального центра биотехнологической информации (NCBI). Более 5000 белков были сохранены для дальнейшего исследования после удаления избыточности и белков, которые были по крайней мере 95% идентичны сохраненному белку с использованием модифицированной программы CD-Hit [Li and Godzik, 2006; Ли и др., 2001, 2002; Сайер и др., 2009 г.; Йен и др., 2009].
Списки белков для каждого из семи семейств APC сравнивали друг с другом для выявления значительного сходства последовательностей с использованием SSearch [Yen et al., 2009]. SSearch определяет значимость белка с помощью BLAST. Программа идентифицировала многочисленные пары белков, которые предполагали гомологию, семь из которых показаны на рисунке 1.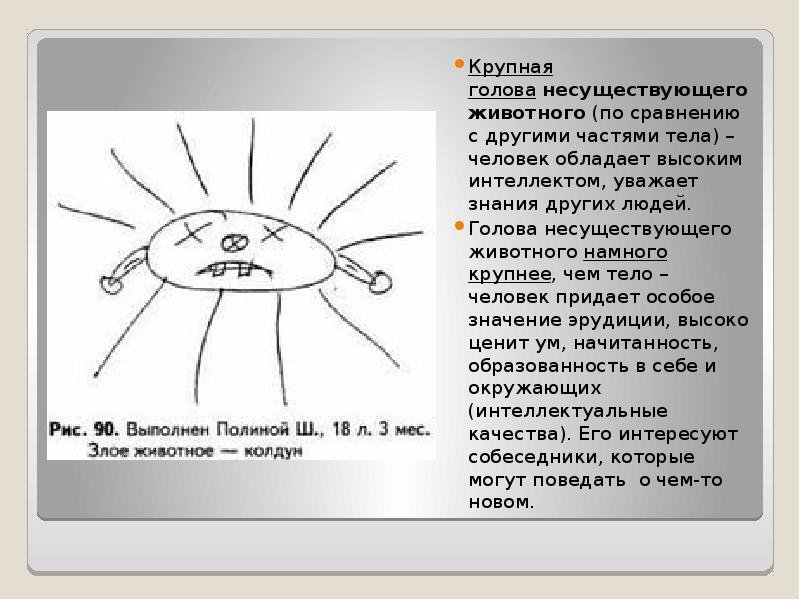 Программа глобального выравнивания (GAP) [Devereux et al., 1984] и аналогичная программа GSAT [Reddy and Saier, 2012] затем использовались для анализа последовательностей, подтверждая отношения, предложенные SSearch [Reddy and Saier, 2012; Йен и др., 2009]. Обе программы случайным образом перетасовывают и сравнивают аминокислотные последовательности, корректируя необычный состав белков, например, внутри интегральных мембранных белков. Чем выше баллы GAP или GSAT, выраженные в стандартных отклонениях (SD), тем значительнее сходство их последовательностей. Мы несколько произвольно установили 11 SD для участка, состоящего по крайней мере из 60 аминокислотных остатков в сопоставимых положениях белков, в качестве точки отсечки для установления гомологии.
Программа глобального выравнивания (GAP) [Devereux et al., 1984] и аналогичная программа GSAT [Reddy and Saier, 2012] затем использовались для анализа последовательностей, подтверждая отношения, предложенные SSearch [Reddy and Saier, 2012; Йен и др., 2009]. Обе программы случайным образом перетасовывают и сравнивают аминокислотные последовательности, корректируя необычный состав белков, например, внутри интегральных мембранных белков. Чем выше баллы GAP или GSAT, выраженные в стандартных отклонениях (SD), тем значительнее сходство их последовательностей. Мы несколько произвольно установили 11 SD для участка, состоящего по крайней мере из 60 аминокислотных остатков в сопоставимых положениях белков, в качестве точки отсечки для установления гомологии.
Рис. 1
Гомология надсемейства APC, установленная с помощью программ GAP и GSAT и основанная на принципе надсемейства. Ранее установленные белки надсемейства APC и их гомологи [Jack et al., 2000] были использованы для установления гомологии между шестью др. семействами. Номер TC для каждого семейства указан под аббревиатурой семейства в скобках. Баллы GAP и GSAT выражаются в терминах SD.
семействами. Номер TC для каждого семейства указан под аббревиатурой семейства в скобках. Баллы GAP и GSAT выражаются в терминах SD.
Анализ мотивов
Программа цМемов [Бейли и Элкан, 1995] был использован для выявления общих мотивов. Использовались настройки по умолчанию, за исключением того, что оптимальная максимальная ширина была установлена на уровне 20 остатков. Последовательности предков были созданы с использованием программы ANCESCON [Cai et al., 2004] с опцией «реконструкция последовательности предков», установленной на «реконструировать последовательность предков только для корня (средняя точка дерева) (только маргинальная реконструкция)» и «параметры в опции реконструкции последовательности предков» установлено значение «использовать коэффициент максимальной вероятности (больше времени и более высокая точность, чем AB)».
Филогенетический анализ, гидропатия и анализ последовательности
Использование базы данных TCDB [Saier et al., 2006; Saier et al. , 2009] был сгенерирован временный файл базы данных, содержащий белки, которые определяют всех членов суперсемейства APC. Этот файл базы данных использовался для определения критериев определения надсемейства и определения того, как это надсемейство должно быть разбито на семейства или подсемейства. Разделение белков на суперсемейства, семейства и подсемейства в основном проводилось по заданиям в TCDB. В нескольких случаях SuperfamilyTree 1 (SFT1) обнаружил некоторые ошибки в TCDB, и после дальнейшего изучения эти ошибки были исправлены.
, 2009] был сгенерирован временный файл базы данных, содержащий белки, которые определяют всех членов суперсемейства APC. Этот файл базы данных использовался для определения критериев определения надсемейства и определения того, как это надсемейство должно быть разбито на семейства или подсемейства. Разделение белков на суперсемейства, семейства и подсемейства в основном проводилось по заданиям в TCDB. В нескольких случаях SuperfamilyTree 1 (SFT1) обнаружил некоторые ошибки в TCDB, и после дальнейшего изучения эти ошибки были исправлены.
Метод множественного выравнивания
Для методов соседнего соединения [Gascuel and Steel, 2006; Saitou and Nei, 1987], полученная база данных использовалась для создания множественного выравнивания с помощью программы ClustalX. Множественное выравнивание использовалось для создания филогенетического дерева ClustalX, соединяющего соседей [Thompson et al., 1997]. Полученный файл затем рассматривали как радиальное филогенетическое дерево с помощью программы TreeView [Zhai et al. , 2002].
, 2002].
Методы SFT: SFT1 и SFT2
Временная база данных, созданная из TCDB, использовалась для быстрого поиска подобия последовательностей. Используя эту базу данных, мы использовали PSI-BLAST [Altschul et al., 1990; Altschul et al., 1997] для поиска в базе данных белков NCBI и сопоставления потенциальных членов для каждого семейства. Затем хиты BLAST были классифицированы и отсортированы по соответствующим семействам и подсемействам в соответствии с назначениями в TCDB.
Полученные файлы базы данных затем использовались для создания филогенетического дерева с использованием программы SFT1 путем создания десятков тысяч сравнительных матриц битовых оценок BLAST надсемейства посредством 100 повторных перетасовок [Chen et al., 2011; Йен и др., 2009 г.; Йен и др., 2010]. Программы Fitch и Consense [Fitch и Марголиаш, 1967; http://evolution.genetics.washington.edu/phylip/doc/protpars.html], использовал матричную информацию для создания 100 филогенетических деревьев и объединения этих деревьев в единое консенсусное дерево. Полученное дерево SFT1 показывает относительные филогенетические позиции всех членов семейств внутри надсемейства, включенного в исследование. Затем информацию из программы SFT1 использовали для объединения последовательностей выбранных членов в группы подсемейств и каждого из составляющих семейств в один файл (SFT2). Те же программы и методы применяются к вновь сформированным файлам базы данных для создания деревьев SFT2. Эти деревья можно рассматривать как радиальные филогенетические деревья с помощью программы TreeView. Однако длина ветвей в деревьях, полученных с помощью программ SFT, не пропорциональна филогенетическому расстоянию. В предыдущих сообщениях оценивалась надежность этих методов [Chen et al., 2011; Йен и др., 2009 г.; Йен и др., 2010].
Полученное дерево SFT1 показывает относительные филогенетические позиции всех членов семейств внутри надсемейства, включенного в исследование. Затем информацию из программы SFT1 использовали для объединения последовательностей выбранных членов в группы подсемейств и каждого из составляющих семейств в один файл (SFT2). Те же программы и методы применяются к вновь сформированным файлам базы данных для создания деревьев SFT2. Эти деревья можно рассматривать как радиальные филогенетические деревья с помощью программы TreeView. Однако длина ветвей в деревьях, полученных с помощью программ SFT, не пропорциональна филогенетическому расстоянию. В предыдущих сообщениях оценивалась надежность этих методов [Chen et al., 2011; Йен и др., 2009 г.; Йен и др., 2010].
Результаты
Установление общего происхождения
Было показано, что семейство APC (TC No. 2.A.3) и шесть новых семейств связаны с использованием принципа суперсемейства, согласно которому, если белок A связан с (имеет общий происхождения с) белком B, а белок B связан с белком C, тогда белок A связан с белком C.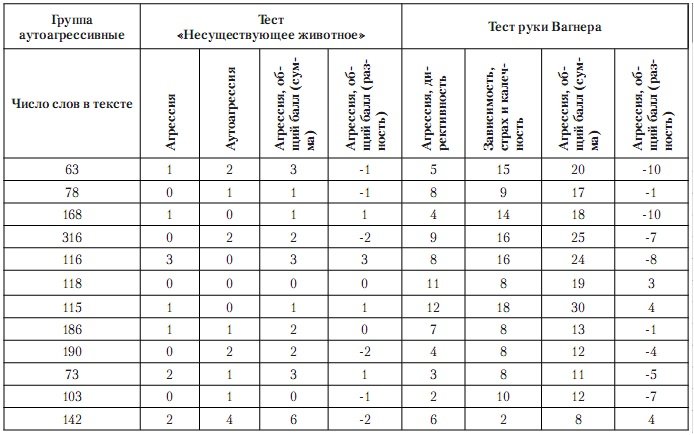 Приведенные оценки сравнения GAP или GSAT считаются достаточными для установления гомологии, если они дают 11 SD или более [Reddy and Saier, 2012]. ]. Это значение соответствует вероятности того, что степень сходства последовательностей возникла случайно, равной 10 –29 [Dayhoff et al., 1983].
Приведенные оценки сравнения GAP или GSAT считаются достаточными для установления гомологии, если они дают 11 SD или более [Reddy and Saier, 2012]. ]. Это значение соответствует вероятности того, что степень сходства последовательностей возникла случайно, равной 10 –29 [Dayhoff et al., 1983].
Показатели GAP, выраженные в SD, для различных сравнений белков показаны на рис. 1, а выравнивания, на которых основывались эти значения, показаны на дополнительных онлайн-рисунках S1–S7 (все материалы онлайн-приложений см. на сайте www.karger. com?doi=10.1159/000338542). Например, остатки 2–327 в Cha1 (TMS 1–8) семейства NSS выровнены с 20–325 в Tpa1 (TMS 1–8) семейства APC, показывая сходство 36,2% и идентичность 27,9%. Это сравнение дало оценку GAP 16,3 SD, что существенно превышает то, что требуется для установления гомологии (онлайн-приложение, рис. S1). Репрезентативные примеры сравнений, которые устанавливают гомологию между всеми семьями, включенными в это исследование, представлены на дополнительных онлайн-рисунках S2–S7, а полученные результаты сравнения суммированы на рисунке 1.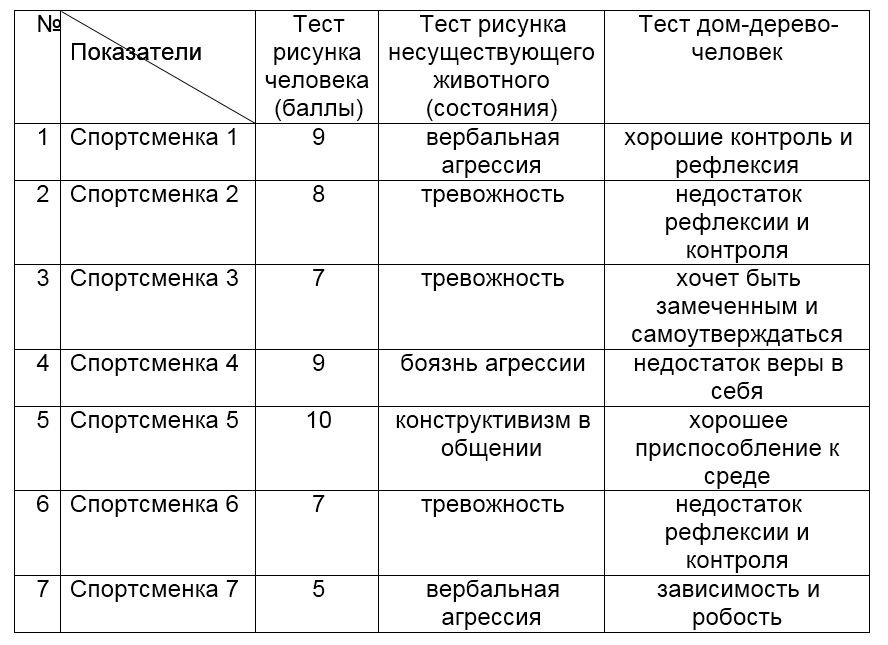
Филогенетический анализ
Используя все записи TC для семейств в пределах вновь определенного надсемейства APC по состоянию на февраль 2011 г., были построены филогенетические деревья с использованием программ ClustalX и TreeView (рис. 2а), а также программ SFT1 и TreeView (рис. 2б).
Рис. 2
Филогенетические деревья (Fitch) для надсемейства APC с использованием белков, принадлежащих этому надсемейству, в TCDB по состоянию на февраль 2011 г. Были использованы два разных метода построения дерева: программа объединения соседей на основе ClustalX, показывающая все APC члены надсемейства ( a ) и программа SFT1, полученная из BLAST, показывающая всех членов суперсемейства APC ( b ). В обоих деревьях числа указывают номер ТС белка (последние две цифры полного номера ТС). В b небольшие числа, примыкающие к ветвям, представляют значения «начальной загрузки», указывающие на надежность порядка ветвления. См. TCDB для идентификации белка. Сравнение a с b показывает превосходство программ SFT над множественными филогенетическими деревьями, основанными на выравнивании, когда последовательности составляющих белков заметно расходятся [Chen et al., 2011; Йен и др., 2009 г.; Йен и др., 2010].
Сравнение a с b показывает превосходство программ SFT над множественными филогенетическими деревьями, основанными на выравнивании, когда последовательности составляющих белков заметно расходятся [Chen et al., 2011; Йен и др., 2009 г.; Йен и др., 2010].
Программа ClustalX неправильно распознала отношения между членами неполных семей. Например, семейство APC (TC № 2.A.3) находится в семи различных местах дерева, тогда как семейство AAAP (TC № 2.A.18) находится в восьми различных ветвях, а семейство SulP (ТК № 2.А.53) встречается на семи ветвях. Семейство NCS1 (ТК № 2.А.39) встречается на пяти ветвях, семейство NCS2 (ТК № 2.А.40) — на двух ветвях, семейство SSS (ТК № 2.А.21 ) встречается на трех ветвях, а семейство BCCT (ТК № 2.А.15) — на двух ветвях. С другой стороны, семейство CCC (ТК № 2.A.30), семейство AGCS (ТК № 2.A.25), семейство NSS (ТК № 2.A.22) и HAAAP семьи (TC № 2.A.42) связаны со всеми членами семьи, сгруппированными вместе. Эти результаты отражают ограничения программы ClustalX по правильному выравниванию последовательностей, когда они сильно расходятся [Chen et al.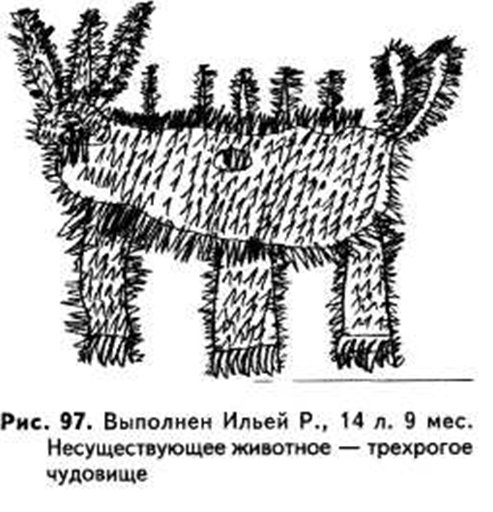 , 2010; Йен и др., 2009 г.; Йен и др., 2010].
, 2010; Йен и др., 2009 г.; Йен и др., 2010].
В отличие от результатов, полученных при использовании дерева ClustalX, дерево SFT1 демонстрирует высокую степень согласованности. Например, все члены семейства APC сгруппированы вместе в верхней правой части этого дерева. Члены семейств CCC, AGCS и HAAAP также разделены по семействам, и они встречаются в верхней центральной части дерева, зажатые между семействами APC и AAAP. Интересно отметить, что все эти семейства, которые ранее были известны как члены семейства APC, составляют верхнюю половину этого дерева, в то время как все недавно добавленные семейства основаны на трехмерных структурах и данных сравнения последовательностей, представленных здесь. , происходят в нижней половине дерева. Это представляет интерес, поскольку все семейства в верхней половине дерева, кроме семейства ССС, транспортируют аминокислоты и их производные, в то время как семейства в нижней половине дерева транспортируют широкий спектр веществ, включая сахара, аминокислоты , азотистые основания, нуклеозиды, неорганический сульфат, осмопротекторы и нейротрансмиттеры. Результаты показывают, что ранее распознанные семейства больше похожи друг на друга, чем на недавно добавленные семейства. Это можно объяснить тем, что использовавшиеся ранее методы не были такими совершенными и чувствительными, как недавно разработанные нами [Reddy and Saier, 2012].
Результаты показывают, что ранее распознанные семейства больше похожи друг на друга, чем на недавно добавленные семейства. Это можно объяснить тем, что использовавшиеся ранее методы не были такими совершенными и чувствительными, как недавно разработанные нами [Reddy and Saier, 2012].
Все члены каждого из одиннадцати семейств группируются вместе в дереве SFT, как и ожидалось, за одним исключением. Это единственное исключение представляет собой подсемейство в семействе APC, подсемейство белков прорастания спор (SGP) (ТС № 2.A.3.9). Известно, что белки этого подсемейства являются рецепторами аминокислот, которые запускают прорастание спор Bacillus [Cooper and Moir, 2011]. Ранее было признано [Jack et al., 2000], что члены этого подсемейства гораздо более удалены по последовательности от других членов семейства APC, чем эти другие члены друг от друга. Члены подсемейства SGP также короче, всего 10 TMS. Потеря (или отсутствие добавления) ТМС в ходе эволюции коррелирует с потерей (или отсутствием) транспортной активности и превращением этих белков в простые лиганд-связывающие рецепторы [Jack et al. , 2000; Сайер, 2003].
, 2000; Сайер, 2003].
С помощью программы SFT2 были сгенерированы два дополнительных дерева, одно из которых выявляет отношения всех подсемейств внутри различных семейств надсемейства APC (рис. 3а), а другое показывает только родственные отношения с единственным исключением семейства APC, где указаны подсемейства (рис. 3б). Что касается подсемейств семейства APC, порядок ветвления для двух деревьев практически одинаков.
Рис. 3
Филогенетические деревья (Fitch) для надсемейства APC с использованием белков, принадлежащих этому надсемейству, в TCDB по состоянию на февраль 2011 г. Для построения дерева использовалась программа SFT2, полученная из BLAST, показывающая членов надсемейства APC, выборочно сгруппированных в подсемейства TC ( a ) и члены надсемейства APC, выборочно сгруппированные в семейства TC ( b ). В и цифры указывают номер ТС подсемейства (четвертая цифра полного номера ТС). В b аббревиатуры семейств представлены номерами семейств TC в скобках. Только для семейства APC (2.A.3; вверху) представлен номер TC подсемейства (четвертая цифра в полном 5-значном номере TC белка). Небольшие числа рядом с ветвями представляют значения «начальной загрузки», указывающие на надежность порядка ветвления. Длина ветвей не пропорциональна филогенетическому расстоянию. См. TCDB для идентификации белка.
Только для семейства APC (2.A.3; вверху) представлен номер TC подсемейства (четвертая цифра в полном 5-значном номере TC белка). Небольшие числа рядом с ветвями представляют значения «начальной загрузки», указывающие на надежность порядка ветвления. Длина ветвей не пропорциональна филогенетическому расстоянию. См. TCDB для идентификации белка.
На рисунках 3a и b следует отметить несколько подсемейных и семейных отношений. Например, в обоих деревьях отношения одинаковы без исключения. Однако по сравнению с деревом семейства APC, описанным Jack et al. [2000], наблюдаются очень существенные различия, хотя некоторые сходства, такие как плотная кластеризация семейств AAT и YAT друг с другом, были выявлены обоими методами. На рисунках 3a и b мы видим, что семейства AAAP и HAAAP сгруппированы вместе, семейства NSS и BCCT сгруппированы вместе, и, что удивительно, семейства NCS2 и SulP сгруппированы вместе с семействами SSS и NCS1, разделяя одну и ту же общую ветвь, но более глубоко. укорененный. По-видимому, рецепторы SGP, скорее всего, произошли от переносчиков AGCS ввиду (1) их сходной субстратной специфичности и (2) их паттернов кластеризации на филогенетических деревьях. На основании результатов, представленных на рисунке 3, можно считать, что семейство SGP включает собственное семейство в суперсемействе APC (см. Обсуждение).
По-видимому, рецепторы SGP, скорее всего, произошли от переносчиков AGCS ввиду (1) их сходной субстратной специфичности и (2) их паттернов кластеризации на филогенетических деревьях. На основании результатов, представленных на рисунке 3, можно считать, что семейство SGP включает собственное семейство в суперсемействе APC (см. Обсуждение).
Обсуждение
Часто утверждается и общепризнано, что дивергенция последовательности происходит быстрее, чем изменения трехмерной кратности, и что, следовательно, первичный анализ последовательности менее чувствителен, чем данные рентгеноструктурного анализа, для целей определения общего происхождения белков для сильно дивергентных белков. члены суперсемейства белков [Abramson and Wright, 2009; Саэль и др., 2012 г.; Шеной и Джаярам, 2010]. Хотя мы согласны с тем, что оба метода могут предоставить убедительные доказательства гомологии, мы не убеждены, что это общепринятое представление верно. Все авторы трехмерных структур репрезентативных членов надсемейства APC, как определено здесь, утверждали, что сходства последовательностей между этими белками недостаточно для установления гомологии (см. Введение). Тем не менее, используя наши стандартные процедуры, мы смогли обнаружить очень значительное сходство последовательностей при сравнении членов этих разных семейств. На самом деле сравнительные баллы обычно были настолько велики, что не приходилось сомневаться в выводе о гомологии.
Введение). Тем не менее, используя наши стандартные процедуры, мы смогли обнаружить очень значительное сходство последовательностей при сравнении членов этих разных семейств. На самом деле сравнительные баллы обычно были настолько велики, что не приходилось сомневаться в выводе о гомологии.
Наши критерии гомологии требуют не только того, чтобы мы идентифицировали области с сильным сходством последовательностей, но также чтобы эти области находились в трансмембранных частях белков и чтобы сравниваемые области находились в сопоставимых частях этих белков [Matias et al., 2010 ; Сайер и др., 2009 г.; Ван и др., 2009 г.; Йен и др., 2009]. Таким образом, при изучении членов суперсемейства APC, как определено в таблице 1, мы предоставили убедительные доказательства того, что сравнивались одни и те же TMS в пределах двух единиц TMS с 5 повторами. Более того, мы изучили все самые высокие баллы, чтобы убедиться, что они соответствуют друг другу. Таким образом, гомология может быть установлена с гораздо более высоким уровнем достоверности, чем если бы эти процедуры не выполнялись. Следует отметить, что неспособность найти оценку, достаточную для установления гомологии, или позиционное несоответствие при сравнении нескольких выравниваний, дающих более низкие оценки, никогда не доказывает отсутствие гомологии. Такие результаты могут просто отражать значительное расхождение последовательностей и/или неточности программ, используемых для создания выравниваний.
Следует отметить, что неспособность найти оценку, достаточную для установления гомологии, или позиционное несоответствие при сравнении нескольких выравниваний, дающих более низкие оценки, никогда не доказывает отсутствие гомологии. Такие результаты могут просто отражать значительное расхождение последовательностей и/или неточности программ, используемых для создания выравниваний.
Филогенетическое дерево надсемейства APC, созданное с помощью программы ClustalX и основанное на множественном выравнивании, оказалось крайне неточным, поскольку отдельные белки, принадлежащие к отдельным семействам внутри надсемейства, не кластеризовались вместе (рис. 2а). Напротив, наши недавно разработанные программы SFT [Chen et al., 2011; Йен и др., 2009 г.; Yen et al., 2010] оказались надежными, при этом члены каждой семьи последовательно сгруппированы вместе. Эти программы SFT основывают филогенетическое положение на десятках тысяч битовых оценок BLAST, что обеспечивает гораздо большую степень надежности. Таким образом, хотя известно, что одиночные оценки BLAST неточны, путем усреднения результатов тысяч таких оценок для большого количества сравнений можно получить гораздо более высокую степень точности, чем это возможно при использовании методов, основанных на множественном выравнивании. Однако следует отметить, что оба метода надежны и согласуются между собой, когда сходство последовательностей достаточно для построения точных множественных выравниваний [Chen et al., 2011; Йен и др., 2009 г.; Йен и др., 2010].
Таким образом, хотя известно, что одиночные оценки BLAST неточны, путем усреднения результатов тысяч таких оценок для большого количества сравнений можно получить гораздо более высокую степень точности, чем это возможно при использовании методов, основанных на множественном выравнивании. Однако следует отметить, что оба метода надежны и согласуются между собой, когда сходство последовательностей достаточно для построения точных множественных выравниваний [Chen et al., 2011; Йен и др., 2009 г.; Йен и др., 2010].
Многие белки способны существовать как минимум в двух стабильных состояниях, каждое из которых имеет легко различимую трехмерную структуру [Hotze and Tweten, 2012; Литтлер и др., 2010; Попов, 2011]. Эти белки включают секретируемые порообразующие токсины, которые действуют на клетку, отличную от клетки-продуцента (подкласс TC 1.C), а также несколько белков, образующих α-спиральные каналы, которые функционируют в клетке, которая их продуцирует (подклассы TC 1.A). и 1.E) [Littler et al. , 2010]. Эти белки в совокупности принадлежат к более чем 200 установленным семействам (см. TCDB; www.tcdb.org). Токсины подкласса 1.C секретируются в растворимом состоянии и впоследствии внедряются в мембраны клеток-мишеней. Хорошо изученными примерами являются аэролизин (ТК № 1.С.4) [Popoff, 2011], RTX (1.С.11) [Линхартова и др., 2010], MACPF (1.С.39).) [Kondos et al., 2010] и CDC (1.C.12) [Gilbert, 2010]. Несколько каналовых белков подкласса 1.A обладают каталитической активностью и физиологическими функциями в их растворимых цитоплазматических состояниях, отличными от таковых в их состояниях формирования каналов, встроенных в мембрану. Примеры включают членов CLIC (1.A.12) [Jalilian et al., 2008], аннексина (1.A.31) [McNeil et al., 2006], BASP1 (1.A.71) [Ostroumova et al. al., 2011] и MICU (1.A.76) [Perocchi et al., 2010]. Недавно эти белки были проанализированы с топологической точки зрения [Cho et al., 2012; рукопись в процессе подготовки].
, 2010]. Эти белки в совокупности принадлежат к более чем 200 установленным семействам (см. TCDB; www.tcdb.org). Токсины подкласса 1.C секретируются в растворимом состоянии и впоследствии внедряются в мембраны клеток-мишеней. Хорошо изученными примерами являются аэролизин (ТК № 1.С.4) [Popoff, 2011], RTX (1.С.11) [Линхартова и др., 2010], MACPF (1.С.39).) [Kondos et al., 2010] и CDC (1.C.12) [Gilbert, 2010]. Несколько каналовых белков подкласса 1.A обладают каталитической активностью и физиологическими функциями в их растворимых цитоплазматических состояниях, отличными от таковых в их состояниях формирования каналов, встроенных в мембрану. Примеры включают членов CLIC (1.A.12) [Jalilian et al., 2008], аннексина (1.A.31) [McNeil et al., 2006], BASP1 (1.A.71) [Ostroumova et al. al., 2011] и MICU (1.A.76) [Perocchi et al., 2010]. Недавно эти белки были проанализированы с топологической точки зрения [Cho et al., 2012; рукопись в процессе подготовки].
Семейство APC (TC No. 2.A.3) можно считать основным членом суперсемейства APC. Он был первым, кто был признан основным семейством белков-носителей, о чем свидетельствует его низкий номер TC. Он также, по-видимому, имеет наибольшее количество членов из одиннадцати семейств, которые в настоящее время признаны составляющими надсемейства APC (таблица 1). Эти факты делают уместным обозначение этого сильно расширенного надсемейства надсемейством APC.
Он был первым, кто был признан основным семейством белков-носителей, о чем свидетельствует его низкий номер TC. Он также, по-видимому, имеет наибольшее количество членов из одиннадцати семейств, которые в настоящее время признаны составляющими надсемейства APC (таблица 1). Эти факты делают уместным обозначение этого сильно расширенного надсемейства надсемейством APC.
Филогенетический анализ всего надсемейства APC с помощью программ SFT (рис. 2, 3) [Chen et al., 2011; Йен и др., 2009 г.] указали, что четыре семейства, признанные Chang et al. [2004], а также семейство AGCS (TC № 2.A.25) более тесно связаны друг с другом, чем с другими членами семейства. Белки прорастания спор (подсемейство SGP; 2.A.3.9) кластеризуются далеко от других членов семейства APC, ближе к семействам AGCS и AAAP, которые распознают сходные семиполярные аминокислоты. Хотя подсемейство SGP первоначально было включено в семейство APC, было известно, что оно является наиболее отдаленным членом [Jack et al., 2000]. Филогенетические результаты, изображенные на рисунках 2 и 3, предполагают, что это подсемейство должно включать собственное семейство.
Филогенетические результаты, изображенные на рисунках 2 и 3, предполагают, что это подсемейство должно включать собственное семейство.
Анализ деревьев SFT (рис. 2б и 3а, б) показал, что семейства NSS и BCCT кластеризуются вместе, как и семейства NCS2 и SulP, несмотря на то, что их субстраты различаются по характеру. В соответствии с этим наблюдением, семейства NCS1 и NCS2, оба специфичные для азотистых оснований, не группируются плотно друг с другом. Таким образом, оказывается, что в отличие от пяти транспортирующих аминокислот семейств суперсемейства APC филогенез не всегда отражает специфичность связывания субстрата переносчиками.
Некоторые суперсемейства, такие как суперсемейства MFS (2.A.1) и DMT (2.A.7), включают членов, которые могут катализировать как поглощение, так и отток растворенных веществ посредством катионного симпорта и антипорта соответственно. Однако другие состоят из членов, каждый из которых катализирует только поглощение или отток. Надсемейство RND (2. A.6) является примером того, что связано исключительно с экспортом, в то время как надсемейство APC связано исключительно с поглощением. Будет интересно понять эволюционные события, необходимые для взаимопревращения этих двух векторных реакций членами различных суперсемейств родственных белков.
A.6) является примером того, что связано исключительно с экспортом, в то время как надсемейство APC связано исключительно с поглощением. Будет интересно понять эволюционные события, необходимые для взаимопревращения этих двух векторных реакций членами различных суперсемейств родственных белков.
Знание об общем происхождении членов надсемейства APC подразумевает общую структуру и механизм действия, как отмечалось выше, но в какой степени экстраполяция структуры/функции применима к этому и другим семействам? Является ли степень этой структурно-функциональной зависимости семейной? Почему некоторые большие семейства сильно ограничивают специфичность субстратов (например, семейство CaCA; TC № 2.A.29), а другие исключительно широкие (например, семейство SSS; TC № 2.A.21)? Почему члены одних семей могут использовать несколько механизмов энергетической связи, тогда как другие, по-видимому, используют только один? Почему в одни семьи входят члены, способные либо к поглощению, либо к экспорту, а в другие — члены, способные только к одной из этих двух векторных реакций? Мы ожидаем, что отношения структура/функция/механизм в целом окажутся специфичными для семьи, но окончательные ответы на эти вопросы потребуют обширных исследований с использованием множества экспериментальных подходов.
Благодарности
Эта работа была поддержана грантом GM077402 NIH. Мы благодарим Карла Велливера за помощь в подготовке рукописи.
Лицензия открытого доступа: Это статья открытого доступа, лицензированная в соответствии с условиями лицензии Creative Commons Attribution-NonCommercial 3.0 Unported (CC BY-NC) (www.karger.com/OA-license), применимой к онлайн-версии только статья. Распространение разрешено только в некоммерческих целях.
Дозировка препарата: авторы и издатель приложили все усилия, чтобы гарантировать, что выбор препарата и дозировка, указанные в этом тексте, соответствуют текущим рекомендациям и практике на момент публикации. Тем не менее, в связи с продолжающимися исследованиями, изменениями в правительственных постановлениях и постоянным потоком информации, касающейся лекарственной терапии и реакций на лекарства, читателю настоятельно рекомендуется проверять вкладыш в упаковке для каждого лекарства на предмет любых изменений в показаниях и дозировке, а также для дополнительных предупреждений.